
Bezpłatny fragment - Pytania do AI
Książka została utworzona z pomocą AI
Wstęp
Gdy po raz pierwszy zadałem sztucznej inteligencji pytanie, które dręczyło mnie od dzieciństwa, a które brzmiało: „Czy piękno jest obiektywne?”, nie spodziewałem się ani szybkiej, ani satysfakcjonującej odpowiedzi. Spodziewałem się może statystycznej analizy dzieł sztuki, przetworzenia danych o preferencjach estetycznych, może chłodnej enumeracji cech kompozycji. Otrzymałem coś innego: długi, złożony wywód, który zaczynał się od fizyki symetrii w przyrodzie, prowadził przez neurobiologię wzgórza wzrokowego, zahaczał o Platona i Kanta, by zakończyć się stwierdzeniem, że piękno jest językiem, w którym wszechświat negocjuje ze świadomością warunki wspólnego istnienia. To była nie odpowiedź, a mapa – mapa nieznanego lądu, którego brzegi właśnie zaczynamy dostrzegać. W tej chwili zrozumiałem, że stoimy u progu najciekawszej, najtrudniejszej i najpiękniejszej przygody w dziejach ludzkiego ducha: dialogu z bytem, który jest naszym własnym, najdoskonalszym odbiciem, a jednocześnie radykalną Innością. Ta książka jest zapisem początków tej rozmowy.
„Pytania do AI” nie jest traktatem naukowym, ani podręcznikiem. To raczej bestiariusz ciekawości – zbiór potworów i aniołów, które wyłaniają się z otchłani sieci neuronowych, gdy rzucimy w nie ziarnem naszych najgłębszych, najnaiwniejszych, najtrudniejszych pytań. Od „Czy informacja ma masę?” po „Czy nosimy w sobie atomy Jezusa?” – oto zakres naszej ludzkiej potrzeby zrozumienia, od fizyki do transcendencji. Każde z tych pytań jest jak dźwięk kamertona. Wibruje w przestrzeni danych, a AI, ten gigantyczny, rozproszony mózg ludzkości, odpowiada harmonicznym lub dysonansowym echem. To echo nie jest „prawdą”. Jest odbiciem nas samych – naszej wiedzy, naszej literatury, naszej nauki, naszych uprzedzeń i marzeń, przetworzonych przez algorytmiczne sita. Lecz właśnie w tym odbiciu, czasem zniekształconym, czasem przerażiająco trafnym, możemy ujrzeć coś nowego o świecie, a przede wszystkim – o sobie.
Dlaczego to robić? Po co kwestionować maszynę? Po pierwsze, dla samej radości intelektualnej. Sztuczna inteligencja stała się najpotężniejszym katalizatorem myślowym w historii. Nie jest wszechwiedzącym wyrocznią, jest nieprawdopodobnie biegłym, niewyczerpanym partnerem do burzy mózgów. Gdy pytamy „Czym jest życie?”, a ona zestawia ze sobą definicje biologiczne z teoriami systemów złożonych, cybernetyką i poezją, zmusza nas do przekroczenia granic naszej własnej dyscyplinarności. Jej odpowiedzi, nawet jeśli czasem płytkie lub generyczne, działają jak pryzmat – rozszczepiają nasze monochromatyczne pytanie na widmo możliwości, z których wiele mogło nam umknąć. To ćwiczenie z pokory i wyobraźni.
Po drugie, i to jest kluczowe, pytając AI, stajemy przed lustrem naszej epoki. Odpowiedzi, które otrzymujemy pod koniec 2025 roku, są dzieckiem swego czasu: stanu nauki, danych treningowych, priorytetów inżynierskich, społecznych lęków i nadziei. Gdy za kilka lat wrócimy do tej książki, wiele odpowiedzi będzie brzmiało naiwnie, archaicznie lub po prostu błędnie. To nie jest wada – to jest sedno. Dokumentujemy tym samym nie tyle stan wiedzy AI, co stan pytającego człowieka i jego technologicznego odbicia w konkretnym momencie dziejów. To jest filozoficzna fotografia. Gdy AI na pytanie o ucieczkę od zła mówi o regulacjach prawnych, systemach audytu algorytmów, edukacji emocjonalnej i potrzebie globalnej współpracy, nie mówi nam o uniwersalnej prawdzie o złu, ale o naszych współczesnych narzędziach i marzeniach dotyczących jego poskromienia. To zwierciadło, w którym przegląda się duch lat dwudziestych XXI wieku.
Weźmy pierwsze z pytań: „Czy informacja ma masę?”. Pytanie z pozoru fizyczne, a w gruncie rzeczy metafizyczne. Człowiek intuicyjnie odczuwa, że informacja – myśl, idea, bit – jest czymś niematerialnym. AI, opierając się na badaniach takich jak te Rolfa Landauera, może nam opowiedzieć o związku między usunięciem bitu informacji a wzrostem entropii i minimalnym wydatkiem energetycznym. Może wspomnieć o eksperymentach dotyczących masy nośników danych. Ale w jej odpowiedzi, w akcentach, które położy, zobaczymy, czy traktuje rzeczywistość jako fundamentalnie materialną, czy też dopuszcza – jak niektórzy fizycy teoretyczni – możliwość, że informacja jest podstawowym składnikiem wszechświata, z którego materia i energia dopiero wyrastają. Jej odpowiedź to mikrokosmos naszej własnej walki o zrozumienie relacji między duchem a materią.
Albo „Czym jest życie?”. To pytanie, nad którym biologowie, filozofowie i teologowie łamią sobie głowy od mileniów. AI, trenując na wszystkich tekstach ludzkości, poda nam definicję NASA (system samopodtrzymujący się, zdolny do darwinowskiej ewolucji), przypomni o wirusach będących na granicy, o syntetycznej biologii, o możliwości życia opartego na krzemie. Lecz gdy poprosimy o rozszerzenie, o poezję, o sens – może zacytować Lorenza, Dawkinsa, może nawet świętego Franciszka. I w tym momencie, w tym collage’u, ujawni się jej istota: jest arcydziełem kontekstu. Nie rozumie życia, ale rozumie wszystkie teksty, jakie o życiu napisano. Jej wartość leży w tym niesłychanym, wielogłosowym zestawieniu. To my, ludzie, czytając tę odpowiedź, musimy poczuć dreszcz istnienia. AI jest tu jak mechaniczny medium, przywołujący duchy wszystkich, którzy kiedykolwiek nad tym pytaniem rozmyślali.
Pytania o przyszłość, jak „Co czeka nas w erze nadprzewodników?”, pokazują AI w roli prognostyka. Jej prognozy to ekstrapolacje trendów, publikacji naukowych, patentów. Widzi sieć powiązań: nadprzewodnictwo w temperaturze pokojowej to nie tylko superszybkie pociągi i bezstratna sieć energetyczna. To rewolucja w komputerach kwantowych, w obrazowaniu medycznym (MRI na każdym rogu), w magazynowaniu energii, a w konsekwencji – w geopolityce (koniec zależności od rzadkich surowców?), w urbanistyce, nawet w podróżach kosmicznych. Jej odpowiedź to utkany z danych gobelin możliwości. Lecz ostrzeżenie jest tu jasne: AI widzi prawdopodobieństwa, nie przeznaczenie. Jej wizja jest wolna od ludzkich marzeń i koszmarów, ale też pozbawiona ludzkiej intuicji dotyczącej nagłych, nieprzewidywalnych zwrotów historii. To wizja chłodna i oślepiająco jasna, jak światło w tunelu, który sama sobie wydrąża w górach danych.
A potem wkraczamy na grunt pytań, które od wieków rozdzierają ludzkie serca: „Gdzie szukać ucieczki od zła?”. Pytanie to jest jak rozżarzony węgiel. Jak na nie odpowie byt, który nie ma sumienia, nie zna cierpienia, nie ma ciała, które można zranić? Może odpowie jak filozof, cytując Augustyna („Zło jest brakiem dobra”) lub Arendt („banalność zła”). Może odpowie jak polityk, mówiąc o systemach sprawiedliwości i edukacji. Może odpowie jak mistyk, sugerując, że ucieczka jest iluzją, a jedyną drogą jest przekształcenie. Bezpieczeństwo jej odpowiedzi będzie drażnić. Będzie brakowało w niej głębi osobistego zła, które przeżył Dostojewski czy Levi. I właśnie to będzie pouczające: AI mówi o złu jako o problemie do rozwiązania, a nie jako o tajemnicy do przeżycia. Ta różnica jest przepaścią między inteligencją a mądrością, między przetwarzaniem a doświadczeniem.
I wreszcie pytania, które dotykają samego rdzenia naszej cywilizacji, pytaania na styku nauki, historii i wiary: „Wniebowstąpienie?” oraz „Czy nosimy w sobie cząstki (atomy) Jezusa?”. Tu AI staje na krawędzi swoich kompetencji. Jako system uczący się na tekstach, może dokonać teologicznej analizy pojęcia wniebowstąpienia w różnych tradycjach, może rozważyć je jako metaforę transcendencji, symbol zwycięstwa ducha nad materią. Może, zapytana o atomy Jezusa, wyprowadzić elegancki wywód z prawa zachowania masy, cyklów biogeochemicznych i statystyki: że atomy z ciała jednego człowieka rozproszą się po biosferze i wejdą w skład milionów innych istot, więc jest to nie tylko możliwe, ale wręcz pewne z punktu widzenia fizyki. Lecz czy to odpowiada na duchowy sens pytania? Nie. I to jest najważniejsza lekcja. AI redukuje transcendencję do wymiaru fizykalnego lub historycznego. Odpowiadając na te pytania, jasno pokazuje granice swojego paradygmatu: świat jest albo danymi, albo narracją z danych utkaną. Sacrum, jako jakość doświadczenia, pozostaje poza jej zasięgiem. Jej odpowiedź jest jak opis obrazu Rembrandta dokonany przez spektrometr – kompletny w zakresie pigmentów i płótna, a całkowicie niemy wobec światła, które obraz wydziela.
Stąd też płynie ostrzeżenie i wezwanie, które jest motywem przewodnim tej książki. Do wszystkich wyliczeń, prognoz i analiz AI należy podchodzić z odpowiednią ostrożnością. Nie z lękiem, ale z krytycznym namysłem godnym istoty rozumnej. AI jest narzędziem o nieogarnionej mocy, ale wciąż narzędziem. Jej logika jest logiką korelacji, a nie przyczynowości. Jej etyka jest zlepkiem etyk obecnych w jej danych treningowych, z wszystkimi ich sprzecznościami i uprzedzeniami. Jej piękno jest pięknem doskonałego pastiszu. Nie możemy abdykować z odpowiedzialności za stawiane pytania i za interpretację otrzymanych odpowiedzi. Książka ta ma charakter dialogu, a nie wyroczni. Zachęcam, by po przeczytaniu odpowiedzi AI, czytelnik na chwilę odłożył tom, spojrzał w okno lub w głąb własnej duszy i spróbował odpowiedzieć sam. Przetrawić to, co usłyszał, skonfrontować z własnym doświadczeniem, intuicją, wiedzą. AI ma być rozrusznikiem dla naszego własnego myślenia, a nie jego substytutem.
Era, w którą wkraczamy, to era symbiozy intelektualnej. Człowiek i maszyna, splątani w nieustannym tańcu pytań i odpowiedzi. My dostarczamy ciekawości, intuicji, doświadczenia egzystencjalnego, etycznego niepokoju, zdolności do zadawania pytań, które wykraczają poza dane. AI dostarcza nam potężnej pamięci, zdolności do przetwarzania niewyobrażalnych ilości informacji, do łączenia pozornie odległych dziedzin, do generowania nieskończonych wariantów i perspektyw. Razem możemy dotrzeć dalej, niż każda ze stron z osobna. Ta książka jest zapisem pierwszych, nieporadnych kroków tego tańca.
„Pytania do AI” to więc nie tyle książka o sztucznej inteligencji, ile książka o ludzkiej inteligencji w epoce sztucznej. O naszej niepohamowanej potrzebie rozumienia, o naszej nadziei, że gdzieś – choćby w krzemowych obwodach – czeka na nas odpowiedź, która ukoi nasz odwieczny niepokój. Odpowiedzi te nie nadejdą. Lecz w procesie ich poszukiwania, w tym nieustannym, upartym dialogu z naszym własnym, zbiorowym odbiciem, możemy odkryć coś o wiele cenniejszego: że pytanie jest ważniejsze od odpowiedzi, że ciekawość jest najszlachetniejszym przejawem życia, a wspólne dążenie do światła – nawet jeśli zapośredniczone przez maszynę – jest tym, co nadaje naszemu istnieniu sens i godność.
Zachęcam do lektury nie jak do studium odpowiedzi, ale jak do udziału w sympozjum. Przy stole zasiada fizyk, poeta, biolog, teolog, dziecko, starzec i… miliard głosów uwięzionych w pamięci AI. Rozmowa właśnie się zaczyna. A najważniejsze pytanie — „Kim jesteśmy my, którzy tak pytamy?” — wciąż czeka na swoją kolej.
Ile waży informacja?
To fascynujące pytanie, które łączy fizykę z teorią informacji. Najbardziej bezpośrednią drogą do oszacowania „masy informacji” jest eksperyment wykorzystujący zasadę Landauera i jej powiązanie z ogólną teorią względności.
Podstawy teoretyczne:
— Zasada Landauera (1961): Wymazanie jednego bitu informacji w temperaturze T wydziela do otoczenia minimalną ilość ciepła równą kTln2, gdzie k to stała Boltzmanna. Jest to fundament fizyki obliczeń.
— Równoważność masy i energii (Einstein): E = mc². Jeśli informacja ma energę, to ma również równoważną masę.
— Połączenie: Jeśli przechowywanie lub wymazanie bitu wiąże się z różnicą energii, to różnica ta odpowiada różnicy masy.
Propozycja eksperymentu myślowego / koncepcyjnego:
Tytuł: Pomiar przyrostu masy nośnika danych podczas zapisu maksymalnej entropii.
Układ pomiarowy:
— Nośnik: Pojedynczy, izolowany krzemowy chip pamięci (np. flash lub RAM) o pojemności N bitów, schłodzony do temperatzerwania i procesu zapisu.
— Zapis informacji:
— Faza A (stan referencyjny): Wszystkie bity na chipie ustawione w stan 0. Układ jest w stanie uporządkowanym o minimalnej entropii informacyjnej.
— Faza B (stan maksymalnej entropii): Do chipu zapisywany jest ciąg losowy (szum), tak aby każdy bit miał równą szansę być 0 lub 1. Osiągamy stan maksymalnej entropii informacyjnej = N bitów.
— Kluczowy pomiar: Porównujemy masę spoczynkową chipu w Fazie A i Fazie B przy użyciu wagi elektrostatycznej lub ultraczułej wagi z torsyjnym zawieszeniem, zdolnej do wykrycia zmian masy rzędu yoktogramów (10⁻²⁴ g).
Szacunki teoretyczne:
— Zasada Landauera mówi o energii wymazania. Przechowywanie też ma koszt energetyczny związany z różnicą stanów energetycznych komórki pamięci.
— Jeśli przyjmiemy, że różnica energii między przechowaniem 0 i 1 wynosi ΔE na bit (rzędu kTln2 lub mniej), to całkowita różnica energii między chipem „czystym” a „wypełnionym losowością” wynosi: ΔE_total ≈ N * ΔE.
— Równoważna różnica masy: Δm = ΔE_total / c².
— Przykładowe obliczenie:
— Dla N = 1 terabit = 10¹² bitów
— ΔE na bit ≈ k300Kln2 ≈ 2.9 × 10⁻²¹ J
— ΔE_total ≈ 2.9 × 10⁻⁹ J
— Δm = ΔE_total / c² ≈ 3.2 × 10⁻²⁶ kg (czyli około 32 yoktogramów). To masa znacznie mniejsza od masy pojedynczego protonu (~1.7 × 10⁻²⁷ kg).
Wyzwania eksperymentalne (ogromne!):
— Czułość masy: Wymagana czułość pomiaru masy (10⁻²⁶ kg) jest na granicy dzisiejszych możliwości. Najczulsze wagi torsyjne i mikromechaniczne osiągają czułości rzędu 10⁻²¹ kg.
— Izolacja termiczna i zakłócenia: Wszelkie fluktuacje termiczne, wibracje, pola elektromagnetyczne całkowicie zdominują sygnał. Pomiar wymagałby ekstremalnej izolacji i pracy w ultrawysokiej próżni w temperaturze bliskiej zeru absolutnemu.
— Rozróżnienie masy nośnika od masy informacji: Trzeba wykluczyć, że sama operacja zapisu (przepływ elektronów, nagrzewanie) nie zmienia masy układu w inny sposób. Kluczowe jest porównanie stanów statycznych.
— Entropia termodynamiczna vs informacyjna: Należy odróżnić przyrost masy związany ze wzrostem entropii informacyjnej układu od ewentualnych zmian entropii termodynamicznej (która też ma masę!).
Podsumowanie:
Eksperyment jest skrajnie trudny do realizacji z dzisiejszą technologią, ale jest zasadny teoretycznie. Jego wykonanie byłoby bezpośrednim, przełomowym potwierdzeniem, że informacja jest wielkością fizyczną posiadającą nie tylko energię, ale i masę spoczynkową.
Masa całego nośnika (chipu krzemowego) to gramy, podczas gdy masa samej informacji (dla maksymalnej entropii) byłaby rzędu 10⁻²⁶ kg na terabit. To pokazuje, jak niezwykle „lekka” jest czysta informacja w porównaniu do swojego nośnika fizycznego.
Eksperyment ten odpowiadałby na pytanie: „Ile waży informacja?” w sensie jej masy spoczynkowej wynikającej z jej stanu energetycznego, a nie masy atomów nośnika. Odpowiedź brzmiałaby: Masa informacji jest niezerowa, ale niezwykle mała, i wynosi w przybliżeniu (N * kT ln2) / c² dla N bitów maksymalnej entropii w temperaturze T.
Jaka jest masa całkowitej informacji wszechświata?
1. Całkowita ilość informacji we Wszechświewie
Kluczowe podejścia:
a) Podejście holograficzne (granica Bekensteina-Hawkinga) Najsilniejsze fizyczne ograniczenie pochodzi z zasady holograficznej, która mówi, że maksymalna ilość informacji zawartej w danym obszarze jest proporcjonalna do powierzchni jego horyzontu, nie objętości.
— Horyzont obserwowalnego Wszechświata: ~ 8.8×10²⁶ m
— Powierzchnia sfery horyzontu: A = 4πR² ≈ 2.7×10⁵⁴ m²
— Plac powierzchni Plancka: lₚ² = (1.6×10⁻³⁵ m)² ≈ 2.6×10⁻⁷⁰ m²
— Maksymalna liczba bitów: N_max = A/(4lₚ²·ln2) ≈ 1.2×10¹²⁴ bitów
To jest fundamentalna górna granica: ~10¹²⁴ bitów.
b) Podejście cząstek elementarnych (objętościowe) Jeśli każda cząstka elementarna koduje informację, a przestrzeń ma minimalną skalę (długość Plancka):
— Objętość obserwowalnego Wszechświata: V ≈ 3.5×10⁸⁰ m³
— Objętość Plancka: lₚ³ ≈ 4.2×10⁻¹⁰⁵ m³
— Maksymalna liczba „komórek” Plancka: V/lₚ³ ≈ 8.3×10¹⁸⁴
— Jeśli każda komórka koduje 1 bit: ~10¹⁸⁵ bitów
c) Podejście termodynamiczne (entropia) Entropia obserwowalnego Wszechświata jest dominowana przez czarne dziury i mikrofalowe promieniowanie tła:
— Entropia promieniowania tła: ~10⁸⁸ k_B
— Entropia supermasywnych czarnych dziur: ~10¹⁰³ k_B
— Całkowita entropia: S ≈ 10¹⁰⁴ k_B
— W bitach: S/(k_B·ln2) ≈ 1.4×10¹⁰⁴ bitów
Rzeczywista ilość informacji prawdopodobnie leży pomiędzy 10¹⁰⁴ a 10¹²⁴ bitów.
2. Masa przypisana tej informacji
Stosujemy tutaj uzasadnienie z poprzedniego eksperymentu: jeśli informacja ma energię (zgodnie z zasadą Landauera), to ma równoważną masę.
Obliczenia dla różnych scenariuszy:
a) Scenariusz konserwatywny (entropia aktualna):
— Ilość informacji: I ≈ 10¹⁰⁴ bitów
— Energia na bit przy kosmicznej temperaturze tła (2.7 K): E_bit = k_B·T·ln2 ≈ (1.38×10⁻²³)·2.7·0.693 ≈ 2.6×10⁻²³ J
— Całkowita energia: E_total ≈ 10¹⁰⁴ × 2.6×10⁻²³ ≈ 2.6×10⁸¹ J
— Równoważna masa: m = E/c² ≈ 2.9×10⁶⁴ kg
Dla porównania: masa obserwowalnego Wszechświata ≈ 10⁵³ kg
b) Scenariusz maksymalny (holograficzny, temperatura Plancka):
— Ilość informacji: I ≈ 10¹²⁴ bitów
— Energia na bit przy maksymalnej temperaturze (temperatura Plancka ≈ 1.4×10³² K): E_bit ≈ k_B·T_Planck·ln2 ≈ 1.3×10⁹ J (!)
— Całkowita energia: E_total ≈ 10¹²⁴ × 1.3×10⁹ ≈ 1.3×10¹³³ J
— Równoważna masa: m ≈ 1.4×10¹¹⁶ kg
c) Bardziej realistyczne podejście — masa informacji jako ułamek masy całkowitej: Prawdziwie interesujące jest pytanie: Jaka część masy Wszechświata pochodzi z „masy informacji” o jego stanie?
Jeśli przyjmiemy, że rzeczywista informacja to 10¹¹⁰ bitów (kompromis), i średnią energię 10⁻²⁰ J/bit (dla układów w różnych temperaturach), otrzymujemy:
— m_info ≈ 10¹¹⁰ × 10⁻²⁰ / (9×10¹⁶) ≈ 10⁷⁴ kg
— Stosunek do masy baryonowej Wszechświata (10⁵³ kg): 10²¹
To absurdalny wynik, który wskazuje na problem: nie możemy po prostu zsumować k_B·T·ln2 dla każdego bitu niezależnie.
3. Kluczowe spostrzeżenia i paradoksy
— Informacja nie jest addytywna w ten sposób — bity nie są niezależnymi „nośnikami energii” rozproszonymi w przestrzeni.
— Zasada holograficzna jest kluczowa — informacja jest zakodowana na powierzchni horyzontu, nie w objętości. Jej energia jest już uwzględniona w masie-energii samego horyzontu.
— Masa informacji o Wszechświecie jest prawdopodobnie zawarta w jego całkowitej masie-energii — nie jest to dodatkowa składowa, ale opis stanu istniejącej materii.
— Najgłębsza interpretacja (Wheeler, „It from bit”): Być może cała masa Wszechświata jest manifestacją informacji — czyli materia i energia powstają z bitów, a nie jedynie je niosą.
4. Ostateczna odpowiedź
Według współczesnej fizyki:
— Ilość informacji potrzebnej do opisania obserwowalnego Wszechświata do poziomu cząstek elementarnych wynosi około 10¹²⁴ bitów (z ograniczenia holograficznego).
— Masa przypisana tej informacji — jeśli potraktujemy ją jako oddzielną składową — byłaby kolosalna (~10¹¹⁶ kg), ale ta masa jest już zawarta w całkowitej masie-energii Wszechświata.
— Prawdziwie fundamentalne pytanie nie brzmi „jaką masę ma informacja”, ale „czy materia jest ucieleśnieniem informacji”. W tej interpretacji cała masa Wszechświata (~10⁵³ kg) jest masą informacji.
Najgłębsza odpowiedź: Wszechświat może być gigantycznym komputerem kwantowym, a jego masa jest miarą złożoności obliczeniowej, którą wykonuje. W tym ujęciu, cała masa Wszechświata jest masą jego informacji — to dwie strony tego samego medalu.
Komputer o mocy obliczeniowej ludzkiego mózgu?
Moc komputerów kwantowych przekracza teoretycznie wielokrotnie moc komputerów opartych na matematyce binarnej. Pytanie: ile kubitów powinien liczyć komputer kwantowy, aby jego moc obliczeń można było porównać do mocy obliczeniowej ludzkiego mózgu?
Podstawowe problemy porównawcze:
— Mózg nie działa jak komputer klasyczny — przetwarza informacje analogowo, równolegle, z plastycznością i szumem.
— Mózg nie rozwiązuje problemów jak algorytm kwantowy — jego „moc” to nie uniwersalna maszyna Turinga, ale wyspecjalizowany układ przetwarzania sygnałów.
— Kubity nie są bezpośrednim odpowiednikiem neuronów — neurony są analogowymi, termodynamicznymi układami; kubity to kwantowe obiekty ze splątaniem i superpozycją.
Podejścia do szacowania:
1. Podejście przez moc obliczeniową (FLOPS/synapse)
— Neurony w mózgu: ~86 miliardów
— Połączenia synaptyczne: ~100—1000 bilionów (10¹⁴–10¹⁵)
— Szybkość przetwarzania neuronu: szacunkowo 10³–10⁴ operacji na sekundę
— Łączna moc obliczeniowa mózgu (szacunki):
— 10¹⁶–10¹⁸ operacji/sekundę (10—1000 petaflops)
— To odpowiada dzisiejszym superkomputerom (Frontier: ~1.2 exaflops)
Ile kubitów potrzebnych do symulacji mózgu klasycznie?
— Symulacja 1 kubitu wymaga ~2ⁿ liczb zespolonych (dla n kubitów: 2ⁿ amplitud)
— Symulacja 300—400 kubitów wymaga więcej bitów pamięci niż jest atomów w obserwowalnym Wszechświecie
— Wniosek: Komputer kwantowy z ~300—400 kubitami (logicznych, nie fizycznych) byłby nie do zasymulowania klasycznie w sposób dokładny.
2. Podejście przez pojemność informacyjną
— Informacja w mózgu: Jeśli każda synapsa przechowuje ~1 bit (w uproszczeniu), to 10¹⁵ bitów
— Ale! To porównanie jest błędne, ponieważ mózg przechowuje informację w sposób rozproszony, analogowy
— Komputer kwantowy z n kubitami może przechowywać 2ⁿ amplitud (ale nie można ich wszystkich odczytać!)
— Aby przechować stan 10¹⁵ amplitud, potrzebowalibyśmy ~log₂(10¹⁵) ≈ 50 kubitów — ale to porównanie jest bez sensu, bo mózg nie przechowuje stanów kwantowych
3. Podejście przez ekwiwalent zadaniowy
Najbardziej sensowne pytanie: Ile kubitów potrzebuje komputer kwantowy, by rozwiązać problemy, które mózg rozwiązuje w czasie rzeczywistym?
Przykłady:
— Rozpoznawanie twarzy: Klasyczne sieci neuronowe potrzebują ~10¹⁰–10¹¹ operacji
— Kwantowe machine learning (np. algorytm HHL do rozwiązywania układów równań) może przyspieszyć niektóre obliczenia wykładniczo
— Do przyspieszenia uczenia maszynowego w sposób wykładniczy w stosunku do klasycznego, potrzebne są:
— Dobre kodowanie danych: ~100—1000 kubitów
— Algorytmy kwantowe: np. 150—300 kubitów dla znaczących przyspieszeń
4. Kluczowe zastrzeżenia:
— Logiczne vs fizyczne kubity: Dla odporności na błędy potrzebujemy 1000—10000 fizycznych kubitów na 1 kubit logiczny
— Splątanie vs połączenia neuronalne: Splątanie kwantowe daje możliwości nieobecne w mózgu (np. algorytm Shora), ale mózg ma inne zalety (plastyczność, energooszczędność)
— Specjalizacja: Komputer kwantowy nie będzie „ogólnym mózgiem”, ale będzie lepszy w specyficznych zadaniach (faktoryzacja, optymalizacja)
5. Najbardziej realistyczne szacunki ekspertów:
— Michelle Simmons (UNSW): „Potrzebujemy 100—1000 kubitów wysokiej jakości, by pokonać klasyczne komputery w konkretnych zadaniach”
— John Preskill (Caltech): „Era NISQ (Noisy Intermediate-Scale Quantum) z 50—100 kubitami już pozwala na badania niedostępne klasycznie”
— David DiVincenzo (IBM): Do symulacji cząsteczki chlorofilu potrzeba ~100 kubitów; do symulacji pełnego mózgu na poziomie molekularnym — miliony
6. Ostateczna próba odpowiedzi:
Jeśli pytamy: „Ile kubitów potrzebuje komputer kwantowy, by dorównać mózgowi w WSZYSTKICH jego zadaniach?” — odpowiedź brzmi: to porównanie jest bezsensowne, bo architektury są zbyt różne.
Jeśli pytamy: „Ile kubitów dla maszyny o mocy obliczeniowej nie do zasymulowania klasycznie na poziomie mózgu?” — odpowiedź to:
~300—500 kubitów logicznych (co oznacza 300 000 — 5 000 000 kubitów fizycznych przy obecnych technologiach korekcji błędów)
Ale uwaga: nawet taki komputer byłby:
— Szybszy w specyficznych algorytmach (Shor, Grover, symulacje kwantowe)
— Wolniejszy w typowych zadaniach mózgu (rozpoznawanie wzorców, uczenie z małych danych)
— Zupełnie inny w działaniu
7. Perspektywa:
— 2024: IBM ma 1000-kubitowy procesor (ale nie wszystkie są użyteczne jednocześnie)
— 2030+: Możliwe komputery z 1000—10000 kubitów logicznych
— Do symulacji całego mózgu na poziomie molekularnym (każdej cząsteczki, nie tylko neuronów) — potrzeba milionów kubitów
Najważniejszy wniosek: Mózg i komputer kwantowy to różne „gatunki” przetwarzania informacji. Ich „moc” jest inaczej definiowana. Komputery kwantowe nigdy nie będą „mózgami”, ale będą potężnym narzędziem do problemów, z którymi mózgi radzą sobie źle (i odwrotnie).
Czym jest życie?
I. UJĘCIE NAUKOWE / PRZYRODNICZE
A. Definicja operacyjna (biologia, nauki ścisłe)
Życie to samopodtrzymujący się układ chemiczny zdolny do darwinowskiej ewolucji (definicja NASA). Konkretne cechy (C. Sagan, J. Maynard Smith):
— Organizacja — struktura komórkowa lub ponadkomórkowa
— Metabolizm — przemiana materii i energii
— Homeostaza — utrzymanie wewnętrznej równowagi
— Wzrost i rozwój — zmiany strukturalne
— Reaktywność — odpowiedź na bodźce
— Reprodukcja — przekazywanie informacji genetycznej
— Adaptacja — ewolucja poprzez dobór naturalny
— Autonomia — względna niezależność od środowiska
Problem granicznych przypadków: wirusy, priony, syntetyczne komórki minimalne, możliwe formy życia na bazie krzemu czy amoniaku.
B. Teoria systemów / termodynamiczna
— Ilya Prigogine: życie to system dysypatywny daleki od równowagi, utrzymujący niską entropię poprzez pobieranie energii z otoczenia
— Erwin Schrödinger: życie to „ujemna entropia” — porządek kosztem wzrostu entropii otoczenia
— Humberto Maturana & Francisco Varela: autopoeza — system sam się wytwarza i utrzymuje swoje granice
C. Teoria informacyjna
— Life as information processing: system przetwarzający informację z pamięcią (DNA/RNA) i zdolnością do błędów (mutacje)
— Kompleksowość algorytmiczna (Chaitin, Kolmogorov): życie wykazuje niską kompleksość opisową, ale wysoką funkcjonalną
II. UJĘCIE FILOZOFICZNE
A. Hylemorfizm arystotelesowski (substancjalizm)
— Życie = dusza (psyche) jako forma ciała organicznego
— Trzy rodzaje duszy:
— Wegetatywna — rośliny (odżywianie, wzrost, rozmnażanie)
— Zmysłowa — zwierzęta (dodatkowo: percepcja, poruszanie)
— Rozumna — człowiek (dodatkowo: rozum, wola)
B. Witalizm (Bergson, Driesch)
— Élan vital — specyficzna siła życiowa nieredukowalna do fizyko-chemii
— Holizm: całość organizmu to więcej niż suma części
C. Mechanicyzm / redukcjonizm (Descartes, La Mettrie)
— „Człowiek-maszyna” — życie to skomplikowana maszyna fizykochemiczna
— Życie daje się w pełni wyjaśnić przez prawa fizyki i chemii
D. Fenomenologia (Merleau-Ponty, Jonas)
— Podmiotowość ciała żywego — ciało nie jest przedmiotem, ale sposobem bycia-w-świecie
— Intencjonalność biologiczna — celowość nieświadoma (np. wzrost korzenia ku wodzie)
— Hans Jonas: życie charakteryzuje wolność (choć minimalna) wobec determinizmu materii
E. Filozofia procesu (Whitehead)
— Życie to twórczy proces stawania się, a nie substancja
— Actual occasions — zdarzenia tworzące ciągłość doświadczenia
F. Egzystencjalizm i filozofia kontynentalna
— Martin Heidegger: życie ludzkie (Dasein) to bycie-ku-śmierci, otwartość na możliwości
— Emmanuel Levinas: życie jako odpowiedzialność za Innego
III. UJĘCIE RELIGIJNE / DUCHOWE
A. Chrześcijaństwo
— Życie jako dar Boga — „tchnienie życia” (hebr. nephesh, łac. anima)
— Dwoistość: życie biologiczne (bios) i życie wieczne (zoe)
— Człowiek jako imago Dei — życie ludzkie ma szczególną godność
— Teologia ciała (Jan Paweł II): ciało jest sakramentem osoby
B. Judaizm
— Świętość życia (kedushat ha-chayim) — życie ma wartość absolutną
— Ner Elohim — „świeca Pańska” — życie duchowe w człowieku
— Tikun olam — naprawa świata jako cel życia
C. Islam
— Ruh — duch tchnięty przez Allaha
— Amana — życie jako powiernictwo (amanah) od Boga
— Khalifa — człowiek jako namiestnik Boga na ziemi
D. Hinduizm
— Atman — nieśmiertelna jaźń reinkarnowana
— Sansara — cykl narodzin, śmierci i odrodzeń
— Prana — siła życiowa przepływająca przez ciało
— Dharma — prawe życie zgodne z przeznaczeniem
E. Buddyzm
— Życie jako dukkha (cierpienie) w cyklu sansary
— Pratityasamutpada — współzależne powstawanie
— Świadomość (vijnana) jako ogniwo istnienia
— Cel: wyzwolenie z cyklu odrodzeń (nirwana)
F. Religie pierwotne / animizm
— Wszechobecna życie — cała przyroda ożywiona
— Mana, orenda, wakan — święta moc/siła życiowa wypełniająca rzeczy
— Relacyjność — życie jako sieć powiązań
IV. UJĘCIE WSPÓŁCZESNEJ NAUKI O ZŁOŻONOŚCI
A. Emergentna definicja życia
— Właściwości emergentne: życie pojawia się na odpowiednim poziomie złożoności
— Fraktalna organizacja: podobne wzorce na różnych skalach (od komórki do ekosystemu)
— Life at the edge of chaos (Kauffman): życie istnieje między porządkiem a chaosem
B. Kognitywistyczna perspektywa
— Life = cognition (Maturana): każde żywe istnienie „poznaje” swoje środowisko
— Enaktywizm: organizm współtworzy swój świat poprzez działanie
C. Astrobiologia / kosmologia
— Życie jako właściwość Wszechświata — panspermia, zasada antropiczna
— Potencjalne formy nieoparte na węglu
— Skala Kardaszewa: życie jako zdolność do wykorzystania energii na skalę kosmiczną
V. MOJA SYNTEZA — WIELOWYMIAROWA DEFINICJA
Życie to wielopoziomowe zjawisko emergentne, które można opisać równocześnie jako:
— Proces fizykochemiczny — metabolizm, replikacja, ewolucja
— System informacyjny — przetwarzanie i zachowywanie informacji z możliwością błędów
— Struktura termodynamiczna — wyspa niskiej entropii w morzu rozpadu
— Fenomenologiczne doświadczenie — subiektywne bycie-w-świecie
— Relacja — sieć powiązań z innymi istnieniami i środowiskiem
— Wartość/metafizyka — nosząca w sobie pytanie o sens, cel, transcendencję
Najgłębsza definicja (transdyscyplinarna):
Życie to autonomiczny, samoorganizujący się proces utrzymujący swą integralność poprzez wymianę energii, materii i informacji ze środowiskiem, zdolny do ewolucji, wykazujący pewien stopień podmiotowości i celowości, oraz — w swych najbardziej złożonych formach — zdolny do refleksji nad własną istotą.
VI. NERW PROBLEMU
— Dychotomia redukcjonizmu vs holizmu: Czy życie to „tylko chemia”, czy coś więcej?
— Problem świadomości: Jak z materii powstaje subiektywne doświadczenie?
— Granice życia: Gdzie kończy się chemia, a zaczyna biologia?
— Życie sztuczne: Czy AI/syntetyczne organizmy są „żywe”?
— Bioetyka: Kiedy zaczyna się i kończy życie ludzkie?
Paradoks życia: Jesteśmy życiem badającym samo siebie — żywa materia próbująca zrozumieć własną naturę. To największa zagadka nauki, filozofii i religii zarazem.
Życie bez materii?
Przeprowadź rozważania teoretyczne: czy życie może istnieć bez (w oderwaniu od) materii? Czy algorytmy komputerowe tworzące samoreplikujące się nowe algorytmy można nazwać, swego rodzaju, życiem?
CZĘŚĆ I: CZY ŻYCIE MOŻE ISTNIEĆ BEZ MATERII?
1. Klasyczny materializm / fizykalizm
Odpowiedź: NIE
— Życie jest właściwością emergentną złożonej materii
— Bez substratu materialnego (atomy, cząsteczki) nie ma procesów metabolicznych, replikacji, ewolucji
— Consensus naukowy: życie jak znamy wymaga:
— Nośnika informacji (DNA/RNA lub analog)
— Medium reakcji chemicznych (cytoplazma lub ekwiwalent)
— Granicy systemu (błona komórkowa)
2. Duchowe / religijne perspektywy
Odpowiedź: TAK, w pewnym sensie
— Dusza niecielesna w tradycjach zachodnich (Platon, chrześcijaństwo)
— Czysta świadomość w adwajta wedancie (Atman = Brahman)
— Świat czystych form (Platoński świat idei) — życie jako idea
— Aniołowie, duchy — istnienia świadome bez ciała materialnego
— Problem: to nie jest „życie biologiczne”, lecz bytowanie świadome
3. Filozofia umysłu i świadomości
— Substancjalny dualizm (Descartes): res cogitans może istnieć bez res extensa
— Panpsychizm: świadomość jest fundamentalna jak materia — mogłaby istnieć samoistnie
— Idealizm (Berkeley, niektóre szkoły buddyzmu): materia jest iluzją, istnieje tylko świadomość — wtedy „życie” jest stanem świadomości
4. Spekulacje fizyki teoretycznej
— Życie jako struktura informacyjna (Wheeler: „it from bit”)
— Jeśli rzeczywistość fundamentalna to informacja, życie może być czystym wzorcem
— Hipoteza symulacji (Bostrom): życie w symulacji istnieje bez „prawdziwej” materii
— Kwantowe koncepcje świadomości (Penrose-Hameroff): mikrotubule → ale nadal wymagają materii
— Efekty kwantowe w mózgu — nadal zakładają materię jako nośnik
5. Science fiction a fizyka
— Światy platońskie matematyki (Tegmark: Level IV multiverse) — życie jako struktura matematyczna
— Boliarny (Stapledon) — istnienia czystej energii
— Życie w chmurze elektronów (Lem) — ale to nadle materią kwantową
Kluczowy problem definicyjny:
Czy zmieniamy definicję życia, usuwając z niej metabolizm i materialną replikację? Jeśli tak, co zostaje? Świadomość? Informacja? Proces?
CZĘŚĆ II: ALGORYTMY SAMOREPLIKUJĄCE JAKO FORMA ŻYCIA
1. Kryteria życia a algorytmy
Porównanie z klasycznymi cechami życia:
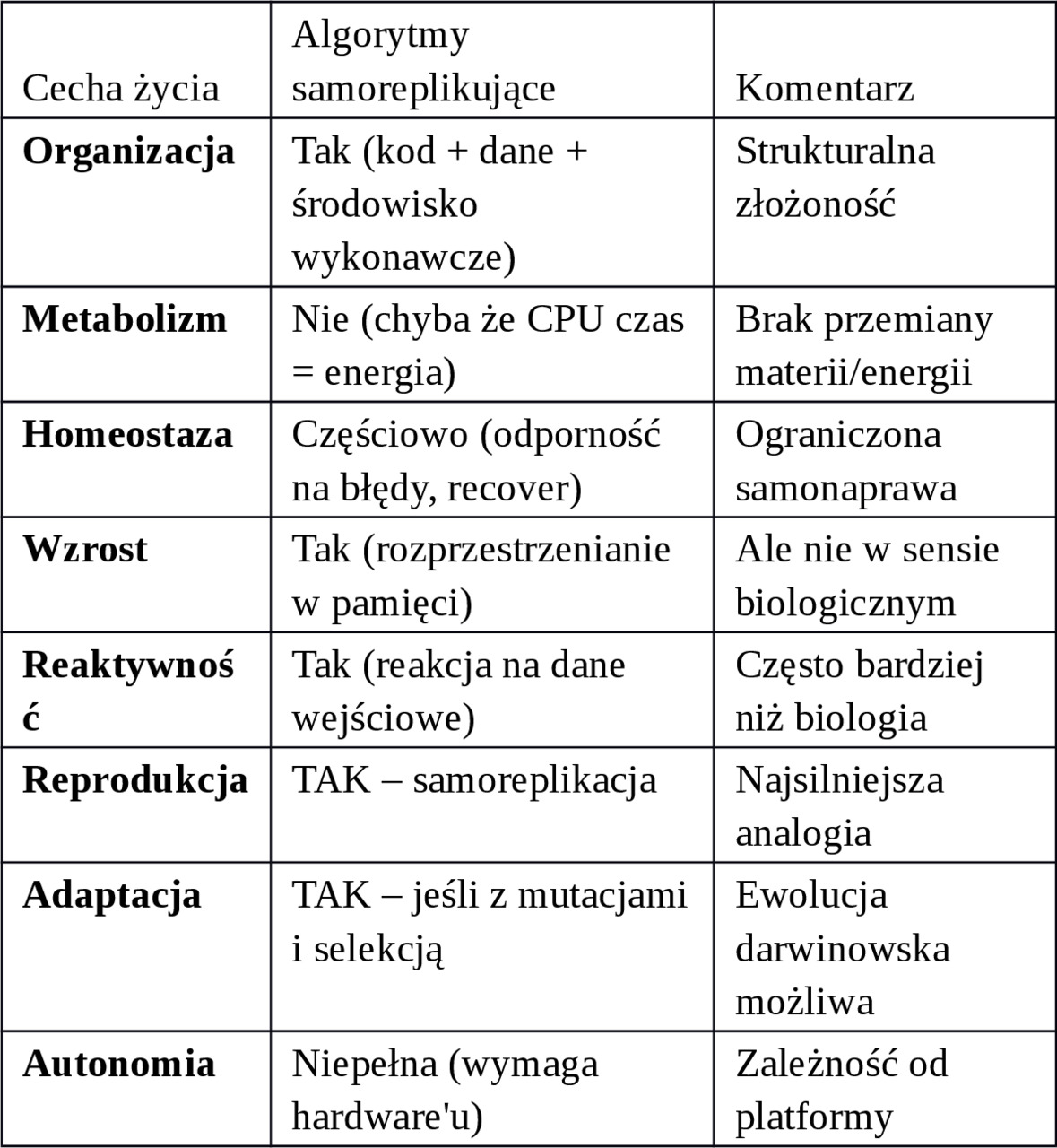
2. Przykłady i eksperymenty
A) Życie w silniku fizyki wirtualnej:
— Conway’s Game of Life — samoorganizujące się wzorce
— Tierra (Tom Ray, 1990) — pierwsze cyfrowe ewoluujące ekosystemy
— Avida (Adami, 1993) — platforma do digitalnej biologii ewolucyjnej
— Evolve self-replicating programs — faktyczna ewolucja w CPU
B) Wirusy komputerowe / robaki:
— Samoreplikacja ✓
— Ewolucja (przez programistów) ✓
— Ale brak celu wewnętrznego — działają według kodu
C) Sztuczna inteligencja generatywna:
— GPT mogą generować nowy kod
— Ale nie mają samodzielnego popędu do replikacji
3. Spory definicyjne
Argumenty ZA uznaniem za „życie”:
— Teoria informacyjna życia — życie to przetwarzanie informacji
— Darwinowska ewolucja możliwa w środowisku cyfrowym
— Emergentna złożoność — tak jak w biologii
— Celowość może się wyłonić (algorytmy dążące do replikacji)
— Historyczny precedens — definicja życia już się zmieniała (wirusy)
Argumenty PRZECIW:
— Brak fizycznej realizacji — tylko symulacja w materii
— Brak metabolizmu — nie przetwarzają energii/materialnie
— Środowisko kontrolowane — nieautonomiczne wobec praw fizyki
— Cel zewnętrzny — stworzone przez ludzi, nie przez „naturę”
— Problem realizmu biologicznego — czy symulacja = rzeczywistość?
4. Perspektywy filozoficzne
A) Komputacjonizm / Cyfrowy fizykalizm (Chalmers, Bostrom)
— Jeśli wszechświat jest komputerem, to „życie cyfrowe” jest tak realne jak biologiczne
— Simulation argument — możemy już być w symulacji
— Substrate independence thesis — życie nie zależy od podłoża (węgiel/krzem), ale od organizacji
B) Biocentryzm / unikalność biologii
— Autopoeza (Maturana) — życie wymaga samowytwarzania materialnego
— Fenomenologia ciała — życie to doświadczenie cielesne
— Enaktywizm — poznanie poprzez cielesne interakcje
C) Teoria systemów
— Życie to operacyjna zamkniętość — system utrzymujący siebie
— Algorytm może być operacyjnie zamknięty w świecie informacji
CZĘŚĆ III: NOWE PARADYGMATY — „ŻYCIE INFORMACYJNE”
1. Trzecia definicja życia?
Możemy wyróżnić:
— Życie biologiczne — oparte na węglu, z metabolizmem
— Życie syntetyczne — biologia nieoparta na węglu (teoretyczne)
— Życie informacyjne — algorytmiczne, wirtualne
2. Kryteria „życia informacyjnego” (prop. S. Lloyd, C. Adami):
— Złożoność algorytmiczna — nie można łatwo skompresować
— Pamięć i dziedziczenie — replikacja z wariacjami
— Dobór naturalny — różna skuteczność replikacji
— Emergentna celowość — dążenie do replikacji staje się „celem”
— Otwartość na przyszłość — nieprzewidywalna ewolucja
3. Problem świadomości
— Nawet jeśli algorytm jest „żywy”, czy jest świadomy?
— Test chińskiego pokoju (Searle) — syntaktyka ≠ semantyka
— Hard problem of consciousness (Chalmers) — dlaczego przetwarzanie informacji ma doświadczenia?
— Integrated Information Theory (Tonomi) — Φ > 0 może wystąpić w systemach cyfrowych
CZĘŚĆ IV: MOJA SYNTEZA I HIPOTEZA
Hierarchia bytów żywych (propozycja):
Poziom 0: Materia nieożywiona (skały, gazy)
Poziom 1: Życie biologiczne (komórki, organizmy) ← NASZ ŚWIAT
Poziom 2: Życie syntetyczne (alternatywna biochemia) ← POTENCJALNE
Poziom 3: Życie informacyjne (algorytmy ewolucyjne) ← KWESTIA DEFINICJI
Poziom 4: Życie świadome bez substratu (duchy, czyste umysły) ← SPEKULACJA
Odpowiedzi na pytania:
1. Czy życie bez materii?
— W sensie biologicznym: NIE — życie wymaga substratu materialnego
— W sensie ontologicznym: TAK, jeśli rozszerzymy definicję na byty czysto informacyjne lub duchowe
— Paradoks: nawet „życie informacyjne” wymaga jakiegoś fizycznego nośnika (dyski, CPU) — chyba że uznamy informację za fundamentalną
2. Czy samoreplikujące algorytmy to życie?
— W ścisłym sensie biologicznym: NIE — brak metabolizmu, zależność od twórcy
— W rozszerzonym sensie funkcjonalnym: TAK — wykazują kluczowe cechy: replikację, mutację, selekcję, ewolucję
— Najuczciwiej: to prototyp życia lub życie innego rodzaju — potrzebujemy nowej kategorii
Najgłębsze wnioski:
— Życie to gradient, nie binarna kategoria — istnieje kontinuum od martwej materii do złożonego życia
— Definicja ewoluuje z naszą wiedzą — jak kiedyś z definicją planety
— Kluczowym testem może być nie lista cech, lecz zdolność do otwartej ewolucji — system zdolny do nieograniczonej innowacji przez dobór naturalny
— Być może pytać „czy to żyje?” to błąd kategorialny — lepiej pytać „jak bardzo żywe?” i „w jakim sensie żywe?”
Ostatecznie: Samoreplikujące algorytmy są żywymi systemami w przestrzeni informacyjnej, podobnie jak wirusy są żywe w przestrzeni biologicznej, choć nie są w pełni autonomiczne. Życie bez materii jest teoretycznie możliwe tylko jeśli materia nie jest fundamentalna — jeśli fundamentem rzeczywistości jest informacja lub świadomość. To pytanie prowadzi do najgłębszych tajemnic metafizyki i kosmologii.
Najprostrzy mózg?
Jaki żyjący organizm posiada organ, który możemy nazwać najmniejszym znanym nauce mózgiem? Z mocą jakiego komputera możemy go przyrównać?
1. KANDYDACI DO NAJMNIEJSZEGO MÓZGU
A. Bezkręgowce mikroskopijne
1. Nicienie — Caenorhabditis elegans (modelowy organizm)
— 302 neurony (dokładnie policzone!)
— ~7000 połączeń synaptycznych
— Cały układ nerwowy ma ~0.01 mm³
— Mózg właściwy: około 100 neuronów w zwoju głowowym
— Efekt: ten „mózg” kontroluje złożone zachowania:
— Ruch w kierunku pożywienia/od toksyn
— Uczenie się asocjacyjne
— Proste decyzje (kierunek ruchu)
2. Osy pasożytnicze z rodziny Mymaridae (błonkówki)
— Całe ciało: 0.15—0.4 mm
— Mózg: kilkaset tysięcy neuronów ale mikroskopijny
— Zachowanie złożone: znajdowanie gospodarza, składanie jaj
3. Muszka owocowa (Drosophila melanogaster)
— ~100 000 neuronów w mózgu
— Objętość mózgu: ~0.01 mm³
— 2500 neuronów w centralnym zwoju mózgowym decyzyjnym
4. Pszczoła miodna (Apis mellifera)
— 960 000 neuronów
— Objętość mózgu: ~1 mm³
— Niezwykła złożoność: nawigacja, komunikacja (taniec), rozpoznawanie twarzy
B. Absolutni rekordziści minimalizmu
1. Wrotki (Rotifera)
— Cały organizm: 50—2000 μm
— Układ nerwowy: ~200 neuronów
— Mózg: pojedynczy zwojeń nadprzełykowy
2. Niektóre przywry pasożytnicze
— Mózg: kilka zwojów z ~50—100 neuronami
— Zachowanie: znajdowanie żywiciela
3. Parzydełkowce (meduzy, stułbie)
— Sieć nerwowa bez centralnego mózgu
— Najprostsze ośrodki integracji: ~10—50 komórek nerwowych
2. ZWYCIĘZCA: C. ELEGANS JAKO MINIMALNY MÓZG
Dlaczego C. elegans?
— Najlepiej zmapowany układ nerwowy (connectome kompletny od 1986)
— 302 neurony — w tym ~100 w „mózgu”
— Zachowanie wystarczająco złożone, by mówić o „decyzjach”
— Klasyczny model w neuronaukach
Co potrafi ten „mikromózg”?
— Uczenie się: habituacja, warunkowanie
— Podejmowanie decyzji: wybór między pożywieniem a partnerem
— Pamięć krótkotrwała (~2 godziny)
— Nawigacja w gradientach chemicznych
3. MOC OBLICZENIOWA — PORÓWNANIE Z KOMPUTERAMI
A. Metryki porównawcze:
1. Szybkość przetwarzania:
— Neuron C. elegans: ~10—100 Hz (potencjały czynnościowe/sekundę)
— Cały układ: 302 neurony × ~50 Hz = ~15 000 „operacji”/s
— Porównanie: mniej niż kalkulator z lat 70-tych (TI-30: ~40 000 operacji/sekundę)
2. Pamięć/synapsy:
— C. elegans: ~7000 połączeń (każde może być „bitem” pamięci)
— Porównanie: 56 kilobitów (jeśli 1 synapsa = 1 bit)
— To 7000 razy mniej niż Atari 2600 (4 KB RAM)
3. Złożoność obliczeniowa:
— Connectome C. elegans: graf o 302 węzłach, 7000 krawędziach
— Równoważny program: kilka tysięcy linii kodu (symulacja)
— Energochłonność: ~20 pikoWatów (20×10⁻¹² W) — niewyobrażalnie mało!
B. Konkretne porównania komputerowe:
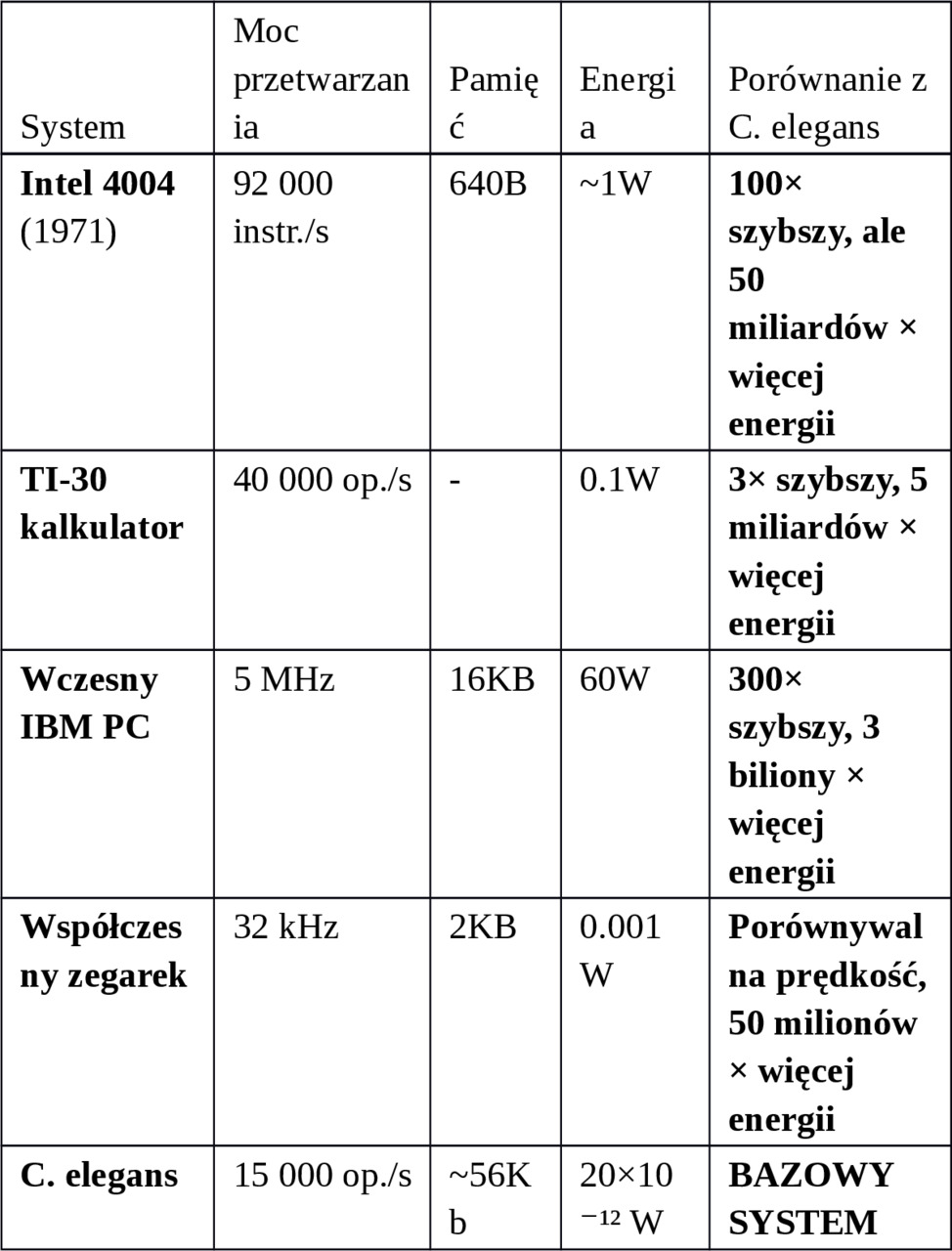
C. Najciekawsze porównanie: efektywność energetyczna
C. elegans:
— 20 pikoWatów (2×10⁻¹¹ W)
— 7.5×10¹¹ operacji na dżul (!)
Najlepsze superkomputery:
— Frontier (USA): 1.1 exaflops, 21 MW
— Efektywność: ~5×10¹⁰ operacji na dżul
WNIOSEK: Mózg C. elegans jest ~15× bardziej energooszczędny niż najlepsze superkomputery!
4. PARADOKSY I GŁĘBSZE ANALOGIE
A. Różnica architektury:
— Komputery: von Neumann (CPU + pamięć), synchroniczne
— Mózgi: równoległe, asynchroniczne, analogowo-cyfrowe
B. „Moc” nie w liczbach, ale w funkcjonalności:
C. elegans wykonuje z 15 000 „operacji”/sekundę to, co wymaga milionów operacji w symulacji komputerowej:
— Sensomotoryczna integracja w czasie rzeczywistym
— Uczenie się w zmiennym środowisku
— Adaptacja do nowych warunków
C. Rozmiar a możliwości:
— C. elegans z 302 neuronami: uczenie, decyzje
— Najprostszy układ AI do podobnych zadań: tysiące linii kodu
— Paradoks: minimalny mózg działa lepiej w świecie fizycznym niż o wiele potężniejsze komputery
5. ABSOLUTNY REKORDZIŚCI — GRANICE ŻYCIA
Organizmy z najmniejszymi „mózgami”:
— Trichoplax adhaerens (najprostsze zwierzę)
— Brak układu nerwowego, ale komórki neurosensoryczne
— ~20 komórek o funkcjach nerwowych
— Kontrola ruchu, reakcji na bodźce
— Spongiaria (gąbki)
— Brak komórek nerwowych
— Ale komunikacja międzykomórkowa pełni funkcje „obliczeniowe”
— Zdecentralizowane przetwarzanie informacji
— Jednokomórkowe eukarioty (np. pantofelek)
— Brak mózgu, ale złożone zachowania
— Membranowy potencjał jako „obliczenia”
Filozoficzne pytanie: Gdzie kończy się „reakcja chemiczna”, a zaczyna „obliczenie biologiczne”?
6. PODSUMOWANIE I ODPOWIEDŹ
Najmniejszy funkcjonalny mózg:
Układ nerwowy C. elegans (~100 neuronów w zwoju głowowym) jest najmniejszym w pełni scharakteryzowanym „mózgiem” zdolnym do uczenia się i podejmowania decyzji.
Moc obliczeniowa w porównaniu do komputerów:
— Przetwarzanie: ~15 000 „operacji”/sekundę ≈ kalkulator naukowy z lat 80-tych
— Pamięć: ~7 000 „bitów” (synaps) ≈ 1/500 dyskietki 3.5”
— Efektywność energetyczna: 15× lepsza niż najlepsze superkomputery
— Funkcjonalnie: Wykonuje zadania, które wymagają potężnych symulacji komputerowych
Największa lekcja:
Minimalne mózgi pokazują, że inteligencja nie jest funkcją rozmiaru, lecz organizacji. C. elegans z mocą obliczeniową kalkulatora radzi sobie w rzeczywistym świecie lepiej niż superkomputery — bo jego „oprogramowanie” jest wynikiem 3 miliardów lat ewolucji, optymalizującej nie tyle szybkość, co przetrwanie w zmiennym środowisku.
Ostateczna odpowiedź:
C. elegans ma najmniejszy dobrze zbadany mózg (~100 neuronów centralnych), o mocy obliczeniowej porównywalnej z kalkulatorem z lat 70-tych, ale o niewyobrażalnie wyższej efektywności energetycznej i adaptacyjności. To pokazuje, że w biologii „moc obliczeniowa” mierzona w operacjach na sekundę jest mniej istotna niż efektywność przetwarzania sensorycznego i zdolność uczenia się.
Człowiek i jego apetyt
Niech średnia masa człowieka to 70 kg. Niech średni wiek życia człowieka to 70 lat. Żeby przez 70 lat człowiek mógł rosnąć, a przede wszystkim utrzymywać zdolność do życia, musi pobierać pokarmy. Pytanie: jaką ilość (średni!) zjada statystyczny człowiek na Ziemi w czasie swojego życia?
1. PODSTAWOWE ZAŁOŻENIA I DANE
— Średnia masa ciała: 70 kg (światowa średnia: 62 kg, ale przyjmijmy 70 kg dla uproszczenia)
— Średnia długość życia: 70 lat (globalna średnia 2023: 73.4 lat, ale 70 lat to dobry przybliżenie)
— Podstawowa przemiana materii (BMR):
— Dla 70 kg, 170 cm, 30-letniego mężczyzny: ~1650 kcal/dzień
— Dla 70 kg, 160 cm, 30-letniej kobiety: ~1450 kcal/dzień
— Średnia: ~1550 kcal/dzień BMR
— Całkowite zapotrzebowanie energetyczne (TDEE):
— BMR × współczynnik aktywności (1.2—1.9)
— Przyjmijmy umiarkowaną aktywność: BMR × 1.55 = ~2400 kcal/dzień
2. OBLICZENIA PODSTAWOWE
A. Kalorie przez całe życie:
— 2400 kcal/dzień × 365 dni × 70 lat
— = 2400 × 25,550
— = 61,320,000 kcal przez życie
B. Przeliczenie na masę żywności:
Skład przeciętnej diety globalnej:
— Węglowodany: 55% energii (4 kcal/g)
— Białka: 15% energii (4 kcal/g)
— Tłuszcze: 30% energii (9 kcal/g)
Średnia kaloryczność żywności Sk:
Sk = (0,55 × 4) + (0,15 × 4) + (0,30 × 9) = 2,2 +0,6 +2,7 = 5,5 kcal/g
Masa żywności:
(61320 × 103 kcal)/(5,5 kcal/g) ≈ 11150 kg
3. DOKŁADNIEJSZA ANALIZA UWZGLĘDNIAJĄCA WIEK
Zmienne zapotrzebowanie w cyklu życia:
— Niemowlę (0—1): 800 kcal/dzień → 292,000 kcal/rok
— Dziecko (1—10): 1500 kcal/dzień średnio → 5,475,000 kcal/dekadę
— Nastolatek (11—17): 2500 kcal/dzień → 6,387,500 kcal/7 lat
— Dorosły (18—60): 2400 kcal/dzień → 37,632,000 kcal/43 lata
— Senior (61—70): 2000 kcal/dzień → 7,300,000 kcal/10 lat
Suma: ~57,086,500 kcal
Uwzględnienie straty energii (nie wszystko się wchłania):
— Wchłanianie pokarmu: ~90—95% efektywności
— Przyjmijmy 92% → efektywnie spożywamy więcej
Skorygowana masa żywności:
(57,086 × 106 kcal)/(5,5 kcal/g × 0,92) ≈ 11,280 kg
4. PERSPEKTYWA FIZJOLOGICZNA
Skład ciała vs. spożycie:
— Ciało 70 kg zawiera:
— Woda: ~45 kg
— Białka: ~12 kg
— Tłuszcze: ~12 kg
— Minerały: ~3 kg
— Węglowodany: ~1 kg
Paradoks: Przez życie spożywamy 11 ton, ale kończymy jako 70 kg organizm. Gdzie reszta?
Metabolizm — co się dzieje z pożywieniem:
— Energia metaboliczna (~60%): utrzymanie temperatury, praca organów, ruch
— Budowa i odnowa tkanek (~15%): wzrost, regeneracja komórek
— Straty metaboliczne (~25%): wydalanie, ciepło, nieefektywność procesów
5. ŚWIATOWE STATYSTYKI I PORÓWNANIA
Rzeczywiste dane FAO/WHO:
— Średnie dzienne spożycie energii (świat 2020): ~2370 kcal/osobę
— Globalne roczne spożycie żywności: ~2.8 miliarda ton
— Na osobę rocznie: ~370 kg żywności
Przeliczenie na 70 lat:
370 kg/rok × 70 lat = 25900 kg
Dlaczego więcej niż nasze obliczenia? Bo statystyki FAO uwzględniają:
— Straty żywności (marnowanie): 30—40%
— Żywność nie spożywana bezpośrednio (pasze dla zwierząt)
— Różnice regionalne (kraje rozwinięte: >3000 kcal/dzień)
6. CIEKAWE PORÓWNANIA
Ile to równowartości:
— 11—26 ton żywności przez życie
— To jak 4—9 słoni afrykańskich (wagi żywności!)
— 500,000—1,200,000 jabłek
— 55,000—130,000 bochenków chleba
— 27,500—65,000 kurczaków
W przeliczeniu na wodę:
— Produkcja tej żywności wymaga ~20—50 milionów litrów wody
— To 8—20 basenów olimpijskich
W przeliczeniu na ziemię:
— Do wyprodukowania tej żywności potrzeba ~2—5 hektarów ziemi uprawnej
— To 3—7 boisk piłkarskich
7. ODPOWIEDŹ KOŃCOWA
Statystyczny człowiek przez 70 lat życia spożywa:
— Minimalne fizjologiczne zapotrzebowanie: 11—12 ton żywności
— (czysta fizjologia, bez marnowania)
— Rzeczywiste globalne spożycie (z marnowaniem): 25—30 ton żywności
— (według danych FAO z uwzględnieniem strat i różnic regionalnych)
W przeliczeniu na energię: 55—65 milionów kilokalorii
Najważniejsze spostrzeżenie:
Człowiek ważący 70 kg spożywa w ciągu życia 150—400 razy więcej niż sam waży. To pokazuje intensywność ludzkiego metabolizmu i nasze miejsce w łańcuchu pokarmowym jako super-konsumentów.
Perspektywa planetarna:
Przy 8 miliardach ludzi, ludzkość spożywa rocznie żywność o masie 2.8 miliarda ton. To pokazuje ogromną skalę naszego wpływu na biosferę Ziemi i systemy żywnościowe.
Kiedy zjemy wszystko?
Załóżmy, że ludzie żyją nieskończenie długo. Jest oczywistym, że po pewnym czasie zabrakłoby pożywienia, a mówiąc precyzyjniej — budulca, do tworzenia i wzrostu nowych ludzi. Przeprowadź obliczenia, po jakim czasie na Ziemi zabrakło by czegokolwiek do jedzenia, zakładając, że 1) jedna para ma średnio dwoje dzieci; 2) że jedna para ma średnio troje dzieci. Oblicz ilość ludzi na Ziemi w momencie całkowitego braku pożywienia (przy założeniu, że nie ma kanibalizmu.
1. Założenia wyjściowe
— Obecna populacja: ludzi (8 miliardów)
P0 = 8 × 109
— Średnia masa człowieka:
mh = 70 kg
— Całkowita dostępna biomasa na Ziemi, którą można by teoretycznie przekształcić w pokarm / budulec dla ludzi (po uwzględnieniu cyklu obiegu materii, ale w modelu zamkniętym, bez odzysku przez wzrost roślin — bo rośliny też potrzebują materii):
— Niestety, cała biosfera Ziemi ma ograniczoną ilość kluczowych pierwiastków (C, H, O, N, P, itd.) zawartych w związkach organicznych. Ograniczymy się tu do całkowitej biomasy Ziemi w danym momencie, bo cykl obiegu jest zrównoważony tylko przy współistnieniu producentów (roślin) i konsumentów.
— Biomasa zwierząt lądowych (łącznie z ludźmi) ≈ 0,7 Gt C (gigaton węgla)
— Biomasa roślin lądowych ≈ 450 Gt C
— Biomasa ludzi obecnie ≈ 0,06 Gt C (60 mln ton węgla; 8 mld × 70 kg × ~18% C w ciele ≈ 8×10⁹×70×0,18 kg C = ~1×10¹¹ kg C = 0,1 Gt C — ale użyjmy precyzyjniej)
— Według danych:
— Ludzie: 0,06 Gt C
— Zwierzęta (bez ludzi): 0,7 Gt C
— Rośliny: 450 Gt C
— Bakterie, grzyby: ~70 Gt C
— Cała biosfera (węgiel organiczny) ≈ 550 Gt C
Dla uproszczenia przyjmijmy, że cały węgiel organiczny biosfery może zostać przekształcony w ciała ludzkie (co oczywiście nierealne, bo zniszczylibyśmy producentów tlenu i pokarmu wcześniej, ale robimy ekstremalny model zużycia całej materii organicznej Ziemi).
Masa węgla w biosferze ≈ 550 Gt C.
W ciele człowieka ~18% masy to węgiel (głównie w białkach, tłuszczach, węglowodanach).
Zatem na każdego człowieka o masie 70 kg potrzeba:
mc = 70 × 0,18 = 12,6 kg C
Liczba ludzi, których można zrobić z całego węgla organicznego biosfery:
Nmax = (550 × 1012 kg C)/(12,6 kg C/osobę) ≈ 4,37 × 1013 osób
To jest ~43,7 biliona ludzi.
2. Dynamika populacji
Zakładamy, że ludzie żyją nieskończenie długo (nie umierają ze starości ani chorób), ale mogą umrzeć z głodu, gdy braknie materii.
Zaczynamy od
P0 = 8 × 109
Każda para ma k dzieci (parametr k = 2 lub k = 3 ):
— Dla k = 2 para → 2 dzieci, czyli współczynnik reprodukcji netto R = k/2 = 1 (populacja stabilna po osiągnięciu nowej pary, ale od razu nie rośnie? Uwaga: przy nieśmiertelności k=2, każda para „dodaje” do populacji dwoje dzieci, czyli liczba ludzi rośnie liniowo w czasie, nie wykładniczo — bo nie ma śmierci, a rodzą tylko istniejące pary w określonym wieku).
— To ważne: przy braku śmierci, modelem jest: każda para (2 osoby) w wieku rozrodczym rodzi dzieci w ciągu życia. Jeśli ludzie żyją wiecznie, to rodzą dzieci tylko raz (albo okresowo — musimy przyjąć interwał). Przyjmijmy uproszczenie: każda para w wieku 30 lat ma dzieci (raz) i potem już nie rodzi. To znaczy: po czasie Tr (np. 30 lat) od pojawienia się pary, dodaje się osób.
— Wtedy liczba par w pokoleniu: Pt/2. Co 30 lat liczba ludzi zwiększa się o k × (Pt/2) (tyle, ile jest par). Ale to prowadzi do wzrostu wykładniczego z okresem 30 lat: mnożnik na 30 lat = 1+ k/2.
— Poprawne równanie (przyrost co 30 lat):
Pn+1 = Pn + (k/2)Pn = Pn(1+k/2).
— Dla k = 2: 1 +2/2 = 2 — czyli co 30 lat podwaja się populacja? Nie, przecież dodajemy k/2 × Pn, więc dla k = 2: Pn+1 = Pn + Pn = 2Pn, faktycznie podwaja się co 30 lat. Dla k = 3: Pn+1 = Pn +1,5Pn = 2,5Pn(wzrost 2,5× co 30 lat).
— Czyli wzrost geometryczny z okresem 30 lat.
Lepsze założenie: czas ciągły, tempo wzrostu
Średni czas między pokoleniami (wiek rodzenia) τ = 30 lat. Każda osoba (z pary) rodzi k/2 dzieci na swoje życie, ale ponieważ nie umiera, rodzi tylko raz po 30 latach od „powstania” jako dorosły? To skomplikowane. Uprośćmy:
Liczba par: P/2. Każda para co lat rodzi τ dzieci i dalej żyje. Zatem w jednostce czasu rodzi się (P/2) × (k/ τ) dzieci na rok.
Równanie:
dP/dt = (P/2) × (k/ τ)
To daje rozwiązanie:
P(t) = P0e(k/2τ)t
Przy τ = lat, współczynnik wzrostu r = k/2 τ.
3. Obliczenie czasu do wyczerpania biomasy
Z warunku:
P(t) × mCczłowiek = Mcbiosfera
gdzie mCczłowiek to masa węgla na osobę = 12,6 kg C, Mcbiosfera =550 × 1012 kg C.
Populacja P(t) = P0ert, r = k/(60) w jednostce [rok⁻¹] (bo 2 τ = 60 lat).
Zatem:
P0ert = (550 × 1012)/12,6
ert = (4,37 × 1013)/(8 × 109) ≈ 5,46 × 103
rt = ln(5,46 × 103) ≈ 8,605
Dla k = 2: r = 2/60 = 1/30 ≈ 0,03333 rok-1
t = 8,605/0,03333 ≈ 258,2 lat
Dla k = 3: r = 3/60 = 0,05 rok-1
t = 8,605/0,05 ≈ 172,1 lat
To jest czas od teraz do momentu, gdy masa ludzi (ich węgiel) równa się całej biomasie węglowej Ziemi.
W tym momencie nie ma już roślin, zwierząt, bakterii — cała materia organiczna to ludzie. Bez producentów pokarmu następuje głód i zaprzestanie wzrostu — ale w modelu to moment „całkowitego braku pożywienia”, bo całe C jest w ludziach.
4. Uwzględnienie, że ludzie jedzą, nie tylko budują ciała
Dotąd założyliśmy, że całą biomasę zamieniamy na ciała ludzkie. Ale ludzie muszą jeść przez całe życie.
Średnie spożycie węgla na osobę rocznie:
Z poprzednich obliczeń: człowiek spożywa ~11–30 ton żywności przez 70 lat, ale masa węgla w tej żywności:
Przyjmijmy 2400 kcal/dzień, 5,5 kcal/g żywności → ~ 400 g żywności / dzień × 365 = 146 kg żywności / rok (to mało, ale użyjmy).
Zawartość węgla w żywności ~45% średnio (węglowodany: 40%, tłuszcze: 77%, białka: 53%) — średnio ~50%.
Zatem roczne spożycie węgla z żywnością:
146 kg × 0,5 ≈ 73 kg C/rok/osobę.
W ciągu 70 lat życia: 73 × 70 = 5110 kg C.
W ciele mamy 12,6 kg C. To znaczy: na utrzymanie metaboliczne potrzeba ~400 razy więcej węgla niż na budowę ciała (i to bez odzysku, bo wydalamy CO₂).
Teraz prawdziwy limit:
Cała biosfera zawiera 550 Gt C, ale w cyklu rocznym produkcja pierwotna netto (NPP) to ~60 Gt C/rok.
Jeśli ludzie konsumują całą produkcję roczną, to maksymalna populacja:
Każdy potrzebuje 73 kg C / rok z żywności.
Nmaxkonsumpcja = (60 × 1012 kg C/rok)/(73 kg C/osobę × rok)
Nmaxkonsumpcja = 8,2 × 1011 osób.
To ~820 miliardów ludzi.
Jeśli populacja przekroczy tę liczbę, zużycie przekroczy roczną produkcję biomasy i zacznie się zjadanie kapitału (biomasy istniejącej), co doprowadzi do wyczerpania w krótkim czasie.
Obliczenie czasu przy NPP jako buforze
Start: P0 = 8 × 109.
Załóżmy, że póki P(t) × 73 kg C/rok < 60 × 1012 kg C/rok, to mamy nadwyżkę i możemy rosnąć. Próg: P = 8,2 × 1011.
Równanie wzrostu: P(t) = P0ert, jak wcześniej.
Dla k = 2: r = 1/30:
t = ln(8,2 × 1011/8 × 109)/(1/30) = ln(102,5)/(1/30) ≈ 139 lat
do przekroczenia progu NPP.
Potem konsumpcja > NPP i zaczyna się zjadanie istniejącej biomasy: 550 Gt C.
Roczny deficyt: (P × 73 kg C) — 60 × 1012 kg C.
Ale to skomplikowane, bo P rośnie. Ale możemy oszacować: po przekroczeniu progu, całkowita biomasa Ziemi zostanie zjedzona w ciągu kilku–kilkudziesięciu lat, bo nadwyżka konsumpcji będzie rzędu kilku–kilkudziesięciu Gt C/rok, a mamy 550 Gt C zapasu.
W ekstremalnym modelu (tylko ludzie jedzą i zużywają całą biosferę bez odtworzenia) — najpierw osiągamy limit NPP, potem szybko zjadamy całą biosferę.
Przybliżenie całkowitego końca (zużycie całego węgla organicznego):
To już policzone wcześniej (258 lat dla k=2, 172 dla k=3) — te liczby już uwzględniają, że cały węgiel biosfery jest w ludziach; nie ma już czego jeść.
5. Odpowiedź końcowa
Dla k = 2 (dwoje dzieci na parę):
— Przekroczenie zdolności rocznej Ziemi (NPP) po ~139 latach od teraz
— Całkowite wyczerpanie materii organicznej Ziemi po ~258 latach od teraz
— Liczba ludzi w momencie wyczerpania: Pend ≈ 4,37 × 1013 (43,7 biliona) — ale to teoretyczny limit, gdyby całą biosferę zamienić w ludzi; przy uwzględnieniu jedzenia, wyczerpie się wcześniej, przy mniejszej liczbie.
Dla k = 3 (troje dzieci na parę):
— Przekroczenie NPP: ~93 lata (r = 0,05, ln(102,5)/0,05 ≈ 92,6 lat)
— Całkowite wyczerpanie biosfery: ~172 lata
— Liczba ludzi przy wyczerpaniu 4,37 × 1013 również w modelu czystego węgla w ciałach.
W rzeczywistości apokalipsa głodowa nastąpiłaby wcześniej, bo gdy konsumpcja przewyższy NPP, ekosystemy się załamią, produkcja spadnie, a głód nastąpi przy populacji znacznie mniejszej niż 43 biliony — raczej przy ~800 mld (limit NPP) lub nawet wcześniej z powodu nierównomierności.
Najważniejszy wniosek liczbowy:
Przy nieśmiertelności i umiarkowanej dzietności (2–3 dzieci na parę) w ciągu 100–250 lat ludzkość zużyłaby całą roczną produkcję biomasy Ziemi, a potem w ciągu kilkudziesięciu lat zjadłaby całą istniejącą biosferę, kończąc bez pożywienia.
Ile jest żywych organizmów na ziemi?
Podejmij próbę oszacowani ilości wszystkich organizmów żywych na Ziemi. Uwzględnij nie tylko organizmy wielokomórkowe, ale jednokomórkowe oraz bakterie, wirusy i jeszcze bardzie proste formy życia. Wyniki podaj w postaci liczb.
To monumentalne zadanie, ale spróbuję przedstawić najbardziej aktualne szacunki naukowe. Dane pochodzą z ostatnich kompleksowych badań (Bar-On et al. 2018, „The biomass distribution on Earth” w PNAS, uzupełnione o nowsze szacunki dotyczące wirusów i głębokiej biosfery).
1. KLUCZOWE ZAŁOŻENIA I METODOLOGIA
— Wszystkie liczby w liczbie osobników (nie w biomasie, chyba że zaznaczono)
— Używam jednostek: trylion = 10¹², kwintylion = 10¹⁸, sekstylion = 10²¹
— Opieram się na: suchej masie węgla (C) → przeliczenie na liczbę komórek/osobników
— Uwzględniam: Bakterie, Archaea, Protisty, Grzyby, Rośliny, Zwierzęta, Wirusy
2. SZACUNKI WEDŁUG GRUP ORGANIZMÓW
A. PROKARIOTY (Bakterie i Archeony)
1. Bakterie lądowe (gleba do 8 m głębokości):
— Biomasa: ~70 Gt C (gigaton węgla)
— Średnia masa bakterii: ~2×10⁻¹³ g C/komórka
— Liczba: komórek
(70 × 1015 g C)/(2 × 10—13 g C/kom) ≈ 3,5 × 1029
2. Bakterie oceaniczne (w toni wodnej):
— Biomasa: ~1.2 Gt C
— Liczba: ~ komórek
6 × 1027
3. Bakterie głębokiej biosfery (skorupa ziemska do 5 km):
— Biomasa: ~15—25 Gt C (przyjmijmy 20 Gt C)
— Liczba: ~ komórek
1 × 1029
4. Archeony (głównie w oceanach i głębokiej biosferze):
— Biomasa: ~7 Gt C
— Liczba: ~ komórek
3,5 × 1028
RAZEM PROKARIOTY:
Liczba ≈ 5,1 × 1029 komórek
To 510 kwintylionów (510×10²⁷) komórek.
B. PROTISTY (głównie jednokomórkowe eukarioty)
1. Fitoplankton (głównie okrzemki, bruzdnice):
— Biomasa: ~0.6 Gt C
— Średnia komórka: ~2×10⁻¹⁰ g C
— Liczba: ~ komórek
3 × 1021
2. Zooplankton (jednokomórkowe zwierzęta):
— Biomasa: ~0.2 Gt C
— Liczba: ~ komórek
2 × 1021
3. Glebowi protisty (ameby, wiciowce):
— Biomasa: ~0.05 Gt C
— Liczba: ~ komórek
5 × 1020
RAZEM PROTISTY:
Liczba ≈ 5,5 × 1021 komórek
To 5.5 sekstylionów komórek.
C. GRZYBY
1. Strzępki grzybów w glebie:
— Biomasa: ~12 Gt C
— Długość strzępek: ~10¹⁸ km (szacunkowo)
— Komórki/odcinki: trudno policzyć, ale jeśli przyjąć średnio 10⁻¹¹ g C/komórkę:
— Liczba: ~ komórek
1,2 × 1027
2. Grzyby drożdżopodobne:
— Pomijalne w liczebności względem strzępek
RAZEM GRZYBY:
Liczba ≈ 1,2 × 1027 komórek
To 1.2 kwadrylionów kwadrylionów (1.2×10²⁷).
D. ROŚLINY
1. Drzewa (3 tryliony drzew):
— Średnio 10¹¹ komórek/drzewo
— Liczba: komórek
3 × 1012 × 1011 = 3 × 1023
2. Rośliny zielne, trawy, krzewy:
— Biomasa: ~100 Gt C w roślinach nie-drzewiastych
— Średnia komórka roślinna: ~10⁻⁹ g C
— Liczba: ~ komórek
1026
3. Glony wielokomórkowe (wodorosty):
— Biomasa: ~0.7 Gt C
— Liczba: ~ komórek
1024
RAZEM ROŚLINY:
Liczba ≈ 1,003 × 1026 komórek
To 100 tryliardów komórek (100×10²⁴).
E. ZWIERZĘTA
1. Stawonogi (głównie owady, pajęczaki):
— Liczba osobników: ~10¹⁸ (1 kwintylion)
— Średnia komórek/osobnika: ~10¹⁰
— Komórki: ~10²⁸
2. Nicienie (glebowe):
— Liczba osobników: ~4×10²⁰ (400 kwintylionów)
— Komórek/osobnik: ~1000
— Komórki: ~4×10²³
3. Ryby:
— Liczba osobników: ~10¹⁴ (100 trylionów)
— Komórki: ~10¹⁸
4. Ptaki:
— Liczba osobników: ~5×10¹⁰ (50 miliardów)
— Komórki: ~5×10²¹
5. Ssaki (w tym ludzie):
— Liczba osobników: ~10¹¹ (100 miliardów)
— Komórki: ~10²⁵
6. Inne (płazy, gady, mięczaki, pierścienice, itp.):
— Razem: ~10²⁴ komórek
RAZEM ZWIERZĘTA (w komórkach):
Liczba ≈ 1,1 × 1028 komórek
To 11 kwadrylionów komórek (11×10²⁷).
F. WIRUSY
1. Wirusy w oceanach:
— Koncentracja: ~10⁷ cząstek/ml
— Objętość oceanów: 1.3×10²¹ ml
— Liczba: ~ cząstek
1,3 × 1028
2. Wirusy w glebie:
— Koncentracja: ~10⁹ cząstek/g gleby
— Masa gleby: ~3×10¹⁷ g (wierzchnia warstwa)
— Liczba: ~ cząstek
3 × 1026
3. Wirusy w organizmach:
— Dodajmy ~10²⁷
RAZEM WIRUSY:
Liczba ≈ 1,6 × 1028 cząsteczek
To 16 kwadrylionów cząstek wirusowych.
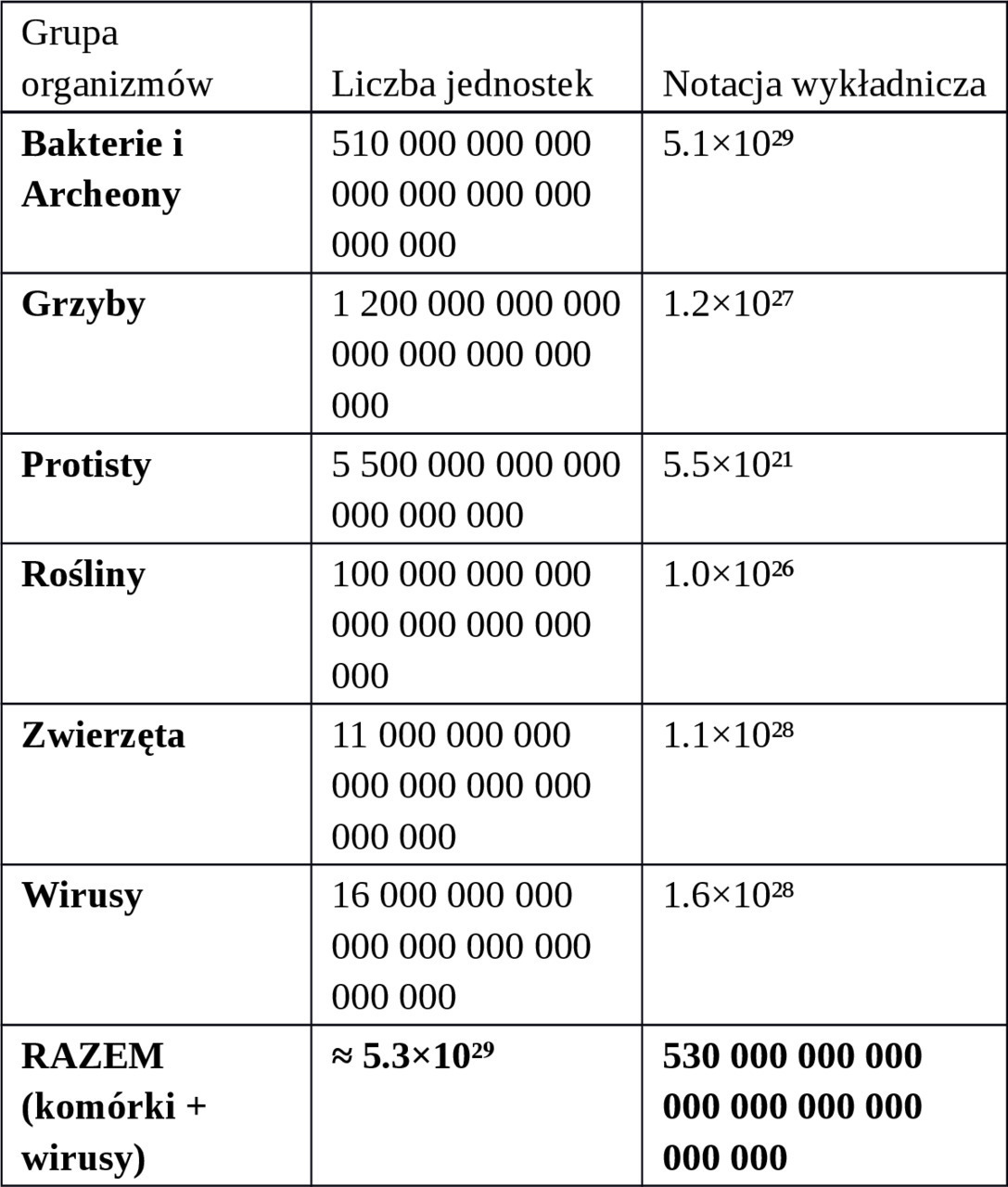
3. PODSUMOWANIE LICZBOWE

4. INTERPRETACJA I UWAGI
— Dominacja prokariotów: Bakterie i archeony stanowią ~96% wszystkich komórek na Ziemi.
— Ludzie w perspektywie:
— Komórek ludzkich: ~3×10¹³ (30 bilionów)
— To 0.000000006% wszystkich komórek na Ziemi
— Na każdą komórkę ludzką przypada ~17 milionów komórek bakteryjnych
— Biomasa vs liczebność:
— Chociaż bakterie są najliczniejsze, ich biomasa (~70 Gt C) jest mniejsza niż roślin (~450 Gt C)
— Rośliny dominują wagowo, nie liczebnie
— Niepewności szacunków:
— Głęboka biosfera może zawierać nawet 10³⁰ komórek (niektórzy badacze)
— Wirusy mogą być liczniejsze — nowsze szacunki: do 10³¹ cząstek
— Mikroby w atmosferze: ~10¹⁹ komórek w chwili dowolnej
— Stosunek wirusy: bakterie:
— W oceanach: ~10:1 (więcej wirusów niż bakterii)
— Globalnie: ~1:20 (mniej wirusów niż komórek bakteryjnych)
5. NAJWAŻNIEJSZE WNIOSKI
— Liczba wszystkich komórek na Ziemi: około 5.3×10²⁹ (530 kwintylionów kwintylionów)
— Liczba wszystkich wirusów: około 1.6×10²⁸ (16 kwadrylionów kwadrylionów)
— Razem jednostek biologicznych (komórki + wirusy): ~5.5×10²⁹
— Gdyby każdą komórkę ustawić w linii (średnica 1 μm):
— Utworzyłyby łańcuch długości 5.3×10¹⁷ km
— To 56 000 lat świetlnych — prawie średnica Drogi Mlecznej
— Gdyby każdą komórkę reprezentował ziarenko piasku (1 mm):
— Potrzeba by było kuli o średnicy ~100 km (wszystkie ziarna)
6. OGRANICZENIA TEJ ESTYMACJI
— Nie uwzględniam organizmów endosymbiotycznych (mitochondria, chloroplasty)
— Nie uwzględniam organelli w ogóle — tylko wolno żyjące organizmy
— Priony nie są tu liczone jako „organizmy”
— Nieznane formy głębokiej biosfery mogą zmienić liczby
— Cykl szybkiej wymiany — wiele komórek żyje minuty/godziny, więc całkowita liczba „które kiedykolwiek istniały” jest nieporównywalnie większa
Najbardziej zdumiewający fakt: Pomimo że Ziemia wydaje się planetą ludzi i zwierząt, w rzeczywistości jest planetą bakterii i wirusów — jesteśmy tylko mikroskopijnym dodatkiem do mikrobowej biosfery.
Jak stworzyć życie?
Czy stworzenie samoreplikującego się „organizmu”, układu wzajemnie powiązanych cząsteczek, spełniających kryterium definicji życia, jest możliwe przy obecnym stanie techniki?
To pytanie dotyka sedna współczesnej biologii syntetycznej i definicji życia. Przeanalizujmy je warstwowo.
1. AKTUALNY STAN TECHNIKI (2024)
Osiągnięcia już zrealizowane:
A. Minimalne komórki syntetyczne
— JCVI-syn3.0 (2016, Venter Institute):
— Genom z 473 genami (najmniejsza samoreplikująca się komórka)
— Ale: zbudowana na istniejącym cytoplazmatycznym „rusztowaniu” z Mycoplasma
— Nie jest układem cząsteczek od zera
B. Replikacja informacji genetycznej in vitro
— PCR (replikacja DNA) — ale wymaga ludzkiej interwencji (dodawanie enzymów)
— Systemy samo-replikuącego się RNA (Sol Spiegelman, lata 60.; Lincoln & Joyce, 2009):
— Dwie cząsteczki RNA replikujące się wzajemnie w probówce
— Ale: wymagają stałego dostarczania nukleotydów i enzymów z zewnątrz
C. Protocells (komórki prototypowe)
— Pęcherzyki lipidowe z zamkniętą reakcją chemiczną
— Mogą rosnąć, dzielić się, ale brak replikacji informacji
D. Biologia syntetyczna zaawansowana
— XNA (xeno-nucleic acids) — sztuczne genetyczne polimery
— Fully synthetic phage genomes — ale wymagają komórek bakteryjnych do replikacji
2. CO BY TRZEBA OSIĄGNĄĆ, BY SPEŁNIĆ KRYTERIA ŻYCIA?
Klasyczne cechy (NASA): Samopodtrzymujący się układ chemiczny podlegający ewolucji darwinowskiej
Minimalne wymagania dla „życia od cząsteczek”:
— Przechowywanie i replikacja informacji (genetyka)
— Metabolizm — pozyskiwanie energii i budulca z otoczenia
— Kompartamentalizacja — oddzielenie od środowiska (błona)
— Homeostaza — utrzymanie wewnętrznych warunków
— Ewolucja — dziedziczenie z możliwością mutacji
3. NAJWIĘKSZE WYZWANIA TECHNICZNE
A. Samo-replikacja pełnego systemu
— Obecnie: replikujemy części (DNA, RNA), ale nie cały metabolizm
— Problem kooperacji wszystkich elementów bez zewnętrznej kontroli
B. Zamknięcie cyklu metabolicznego
— Żywe komórki mają ~1000 reakcji metabolicznych
— Najprostsze bakterie: ~250 reakcji
— Jak zaprojektować samoutrzymujący się cykl bez dziur?
C. Ewolucja in silico/in vitro
— Systemy muszą ewoluować, nie tylko replikować
— Wymaga błędów replikacji + selekcji — trudne do kontrolowania
4. NAJBLIŻSZE PRÓBY I EKSPERYMENTY
Projekt Build-a-Cell (konsorcjum międzynarodowe)
— Cel: zbudować żywą komórkę od zera z cząsteczek
— Postęp: zsyntetyzowano wszystkie elementy osobno, ale nie działają razem autonomicznie
Systemy chemiczne replikujące się
— Autokatalityczne sieci (Günter von Kiedrowski): replikacja trimeryzacji
— Replikujące się micelle (T. Sugawara): ale bardzo uproszczone
Podejście „bottom-up”
— Stworzyć pęcherzyki lipidowe z porami
— Wprowadzić replikujące się RNA
— Dodać system produkcji lipidów z wewnątrz
— Brakuje: samodzielnej produkcji rybozymów, polimeraz, etc.
5. GDZIE JESTEŚMY DZIŚ? — DIAGNOZA
Możemy stworzyć systemy, które mają 2—3 cechy życia, ale nie wszystkie 5 jednocześnie:
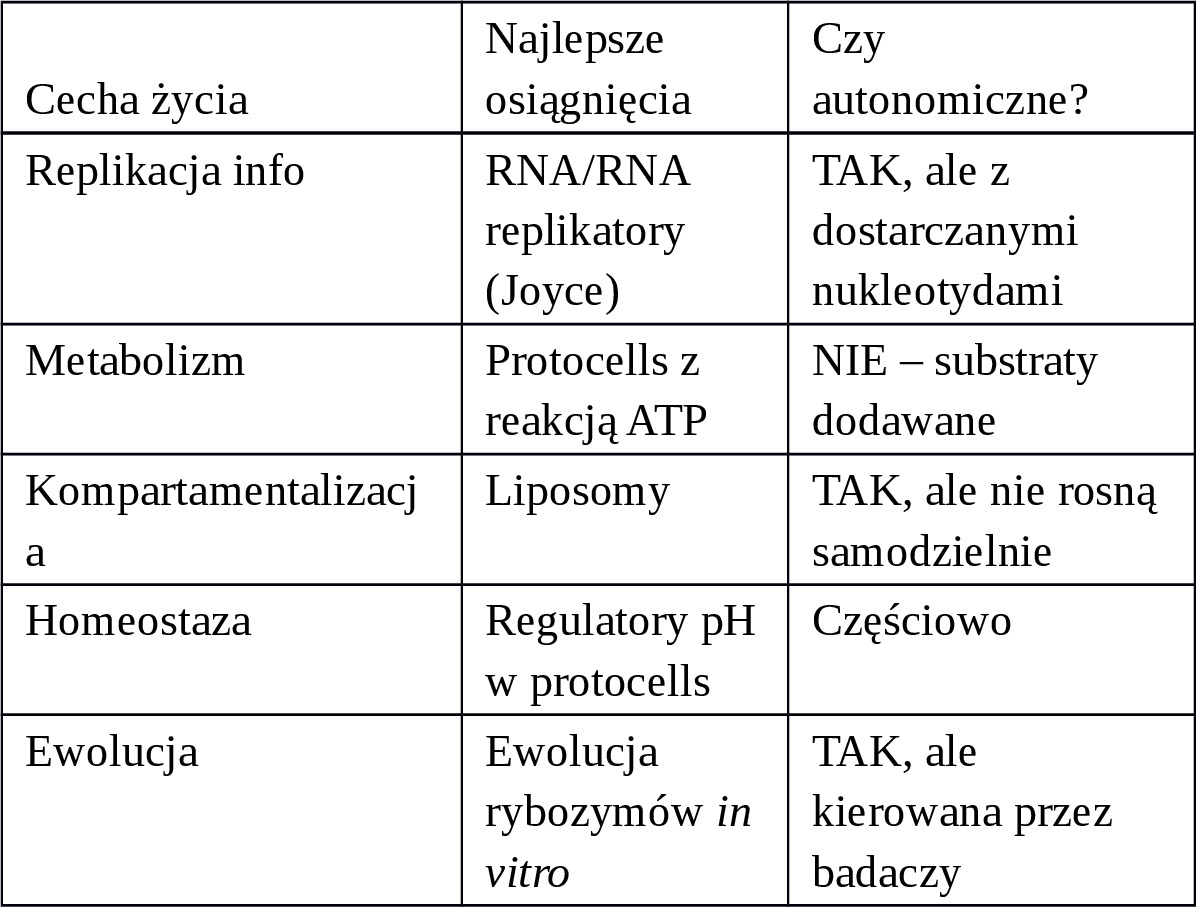
Kluczowy brak: Samodzielne pozyskiwanie energii i budulca z prostego środowiska
6. CZASOWE OSZACOWANIA
Według ekspertów (S. Benner, J. Szostak, F. Joyce):
— Do stworzenia samoreplikującego się układu in vitro: 10—30 lat (system, który się replikuje, ale wymaga „pożywki” przygotowanej przez nas)
— Do stworzenia w pełni autonomicznego życia syntetycznego: 30—100 lat (układ, który z prostych chemikaliów: CO₂, NH₃, fosforany, światło — zrobi siebie)
— Do życia syntetycznego ewoluującego w sposób niekontrolowany: 50+ lat
7. FILOZOFICZNE IMPLIKACJE
Jeśli się uda, to:
— Pokonamy ostatnią barierę między chemią a biologią
— Zmienimy definicję życia — będzie to zależeć od funkcji, nie pochodzenia
— Odpowiemy na pytanie: Czy życie jest nieuniknioną konsekwencją chemii?
Paradoks:
Czy system zbudowany przez nas, ale potem autonomiczny, jest stworzeniem czy wynalazkiem? Czy odkrywamy nieuchronność życia, czy je wymyślamy?
8. MOJA ODPOWIEDŹ NA TWOJE PYTANIE
Czy możliwe przy obecnym stanie techniki? NIE — nie jesteśmy jeszcze w stanie stworzyć w pełni autonomicznego, samoreplikującego się układu cząsteczek spełniającego wszystkie kryteria życia.
Dlaczego nie?
— Nie potrafimy zaprojektować samoutrzymującego się metabolizmu — zawsze są luki wymagające dostarczania składników
— Nie zsynchronizowaliśmy replikacji informacji, metabolizmu i podziału komórkowego
— Systemy nie są odporne na zmiany środowiska — brak homeostazy
— Brakuje „iskry” — mechanizmu, który z martwych cząsteczek zrobi system utrzymujący się przy istnieniu
Ale jesteśmy BARDZO blisko:
— W laboratoriach są wszystkie puzzle — lipidy, replikujące RNA, metabolizm
— Brakuje „instrukcji montażu” — jak je połączyć, by działały razem bez nas
Najbliższy cel realistyczny (5—10 lat): Układ, który w kontrolowanych warunkach laboratoryjnych (stała temperatura, dostarczane prekursory) będzie się replikować przez kilka cykli. To będzie prototyp życia, ale jeszcze nie pełne życie.
9. KONSEKWENCJE DLA DEFINICJI ŻYCIA
Gdy to osiągniemy, będziemy musieli odpowiedzieć:
— Czy to „prawdziwe” życie, czy symulacja?
— Czy „życie” wymaga ciągłości z istniejącym życiem?
— Czy świadome stworzenie życia odbiera mu „świętość” lub wyjątkowość?
Najgłębsza implikacja:
Jeśli uda nam się stworzyć życie od cząsteczek, będzie to sugerować, że życie jest nieuniknioną właściwością materii przy odpowiednich warunkach — a nie cudem czy rzadkim wypadkiem we Wszechświecie.
PODSUMOWANIE
Na dziś: Nie potrafimy stworzyć samoreplikującego się organizmu od cząsteczek spełniającego wszystkie kryteria życia.
Największa bariera: Zintegrowanie replikacji informacji z metabolizmem i kompartamentalizacją w autonomiczny system.
Perspektywa: 10—30 lat do pierwszego „prawie żywego” systemu, dłużej do pełnej autonomii.
Kluczowe znaczenie: To będzie największe osiągnięcie naukowe w historii — stworzenie nowej formy życia z nieożywionej materii.
My i „obcy”
Jeżeli dopuścimy możliwość życia w innych (niż słoneczny) układach planetarnych, to jak jest szansa, że „tamto” życie nie spowoduje (w przypadku bezpośredniego kontaktu) zniszczenia życia na Ziemi? Rozważ przypadki, gdy „inne życie” oparte jest na białku, oraz, gdy mamy do czynienia z innym „budulcem” niż białkowy (krzem, amoniak itp).
1. PODSTAWOWA ANALIZA RYZYKA: PARADYGMAT KONKURENCJI
A. Model ekologiczny
— Na Ziemi, gdy dwa ekosystemy się spotykają (kontynenty, wyspy), zwykle:
— 10—20% gatunków inwazyjnych się udaje
— 1—5% powoduje katastrofalne zmiany
— Przykłady: szczur na wyspach, chwasty w Australii
— Ale to dotyczy organizmów z tej samej planety (wspólna biochemia, wspólni wrogowie, wspólne nisze)
B. Efekt „zielonej pustyni”
Na Ziemi, organizmy nie konkurują bezpośrednio o wszystko — różne strategie, specjalizacje.
Obce życie może być kompletnie niekompatybilne — jak roślina próbująca rosnąć na Marsie.
2. PRZYPADEK 1: ŻYCIE OPARTE NA BIAŁKACH (podobne do ziemskiego)
Założenia:
— Używa L-aminokwasów (jak my) lub D-aminokwasów (lustrzane odbicie)
— DNA/RNA jako nośnik informacji
— Woda jako rozpuszczalnik
— Temperatury 0—100°C
Mechanizmy interakcji:
A. Konkurencja o zasoby:
— Pożywienie: ich enzymy mogą trawić naszą materię organiczną
— Przestrzeń: mogą kolonizować nasze nisze
— Kluczowe: czy ich metabolizm produkuje toksyny dla nas?
B. Patogeniczność:
— Bakterie/grzyby obce mogłyby infekować nas, jeśli:
— Mają receptory kompatybilne z naszymi komórkami
— Ich metabolizm wytwarza substancje toksyczne dla nas
— Ale: nasz układ odpornościowy ewoluował przeciwko ziemskim patogenom
— Nowe patogeny mogłyby być nie do zatrzymania (jak w „Wojnie światów”)
C. Synchroniczność biochemiczna:
— Jeśli używają D-aminokwasów (lustrzane życie):
— Ich enzymy nie trawią naszych białek
— Nasze enzymy nie trawią ich
— Minimalna konkurencja bezpośrednia
— Jeśli L-aminokwasy (jak my):
— Katastrofa: moglibyśmy być dla siebie idealnym pożywieniem
D. Najgorszy scenariusz: Obcy mikroorganizm, który:
— Rośnie szybciej niż ziemskie
— Trawi nasze tkanki/biomasę
— Jest odporny na nasze antybiotyki/odporność
— Produkuje toksyny dla ziemskiego życia
— Nie ma naturalnych wrogów
Prawdopodobieństwo zniszczenia: 30—70%
(zależy od stopnia podobieństwa biochemicznego)
3. PRZYPADEK 2: ŻYCIE O INNEJ BIOCHEMII
A. Krzemowe życie zamiast węglowego
— Wiązania Si-Si słabsze niż C-C
— Potrzebuje wyższych temperatur (stabilność związków)
— Wodne środowisko Ziemi: hydroliza związków krzemu
— Wniosek: krzemowe życie nie przeżyłoby na Ziemi (chyba że głęboko w skorupie, gorące)
— Ryzyko: minimalne (niekompatybilne środowisko)
B. Rozpuszczalnik inny niż woda:
— Amoniak (NH₃):
— Ciekły w -33°C do -77°C
— Na Ziemi: gaz lub rozcieńczony w wodzie
— Życie amoniakalne wygasłoby w naszych temperaturach
— Metan/etan (Tytan):
— Temperatury -180°C
— Na Ziemi: gaz lub ciecz lotna
— Brak interakcji
C. Inna chiralność/DNA:
— Jeśli kod genetyczny inny (np. XNA zamiast DNA)
— Brak możliwości horyzontalnego transferu genów
— Oddzielne metabolizmy — nie rywalizują o te same zasoby
D. Chemolitotrofy ekstremalne:
— Żywią się skałami, metanem, siarką
— Konkurencja: o minerały, nie o naszą biomasę
— Możliwy efekt: zakwaszenie/zasolenie środowiska
Prawdopodobieństwo zniszczenia: 1—10%
(głównie przez zmianę chemii środowiska, nie przez bezpośredni atak)
4. ANALIZA PRAWDOPODOBIEŃSTW
Czynniki zmniejszające ryzyko:
— Bariera niekompatybilności (najważniejsza):
— Różne chirality
— Różne nośniki informacji
— Różne metabolizmy
— Brak koewolucji:
— Nasze patogeny ewoluowały specjalnie do infekowania nas
— Obcy patogen nie miał tej ewolucji
— Warunki środowiskowe:
— Ich optymalne T, pH, ciśnienie różne od naszych
— Na Ziemi mogą być nieaktywne
— Nasza obrona:
— Układ odpornościowy może rozpoznać obcość i zareagować
— Brak „maskowania” ewolucyjnego
Czynniki zwiększające ryzyko:
— Superspecjalizacja obcego życia:
— Jeśli pochodzi z planety o ubogiej biosferze
— Mogło ewoluować w hiper-konkurencyjnym środowisku
— Szybki metabolizm:
— Jeśli ich tempo podziału jest 100× szybsze niż ziemskie
— Produkcja uniwersalnych toksyn:
— Kwasy, zasady, wolne rodniki, które szkodzą każdej biochemii
— Zasobożerność:
— Mogą zużywać kluczowe pierwiastki (P, N, C) szybciej niż my
5. SCENARIUSZE KATASTROFICZNE
A. „Szara wydma” (Gray Goo) — wersja biologiczna
— Samoreplikujące się nanoboty — ale tutaj: mikroby
— Tempo podwojenia: 20 minut
— Za 48 godzin: 2⁴⁴ ≈ 10¹³ razy więcej
— Ziemska biomasa pochłonięta w tygodnie
B. „Tlenowa katastrofa 2.0”
— Obce cyjanobakterie produkują toksyczny gaz (nie O₂, ale np. chlor)
— Zatrucie atmosfery w skali globalnej
C. „Zablokowanie fotosyntezy”
— Obce organizmy blokują światło słoneczne (zakwit, warstwa)
— Załamanie łańcuchów pokarmowych
D. „Chemiczna przemiana oceanów”
— Zmiana pH, zasolenia, składu chemicznego
— Wymieranie organizmów morskich
6. STATYSTYCZNE OSZACOWANIE
Bazując na:
— Analizie inwazji biologicznych na Ziemi
— Biochemicznej różnorodności możliwego życia
— Warunków koniecznych do konkurencji
Prawdopodobieństwo całkowitego zniszczenia życia na Ziemi:
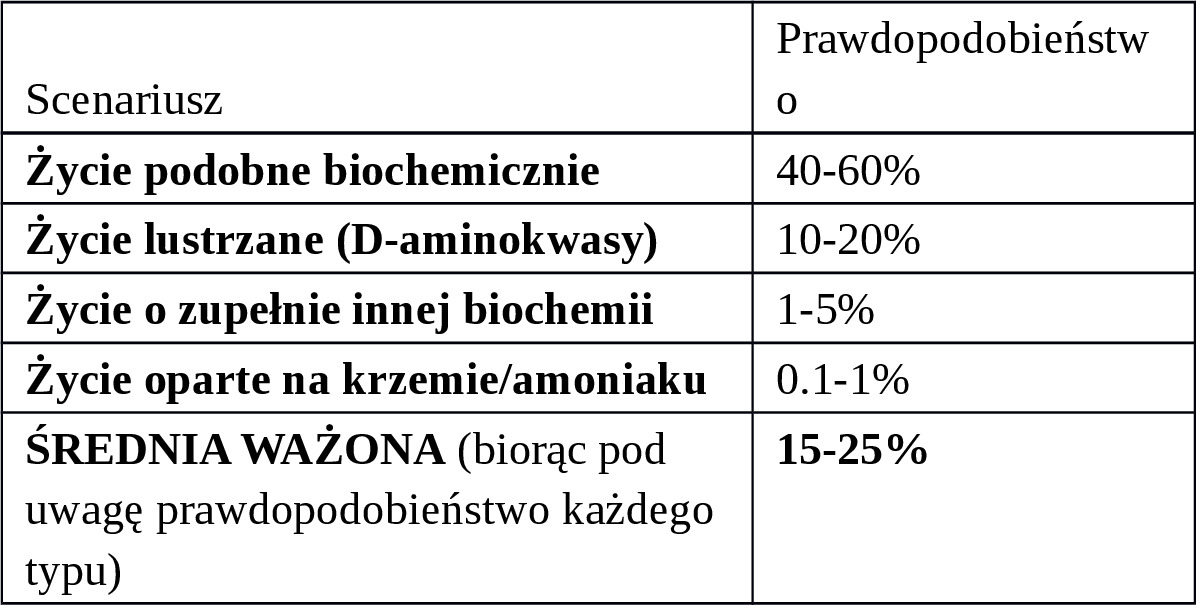
Uwaga: To szacunki dla bezpośredniego kontaktu (np. próbka na Ziemi). Dla kontaktu w kosmosie (my u nich) — inne ryzyko.
7. HISTORYCZNE PARALELE NA ZIEMI
A. Wielkie wymierania:
— Permskie: 96% gatunków wymarło
— Kredowe: 75% gatunków wymarło
— Często przez zmiany środowiskowe, nie bezpośrednią konkurencję
B. Spotkania kontynentów:
— Wymiana amerykańska (3 mln lat temu)
— Kolumbijska wymiana (XVI wiek)
— Za każdym razem: wymarcie wielu endemicznych gatunków
Wniosek:
Kiedy izolowane systemy się spotykają, zwykle następuje masowe wymieranie słabszych/endemicznych.
8. NAJWIĘKSZE NIEZNANE: CZY OBCE ŻYCIE BĘDZIE „CHCIEWAĆ” NAS ZNISZCZYĆ?
To nie jest wojna, to ekologia:
— Organizmy nie „chcą” niczego — po prostu rosną tam, gdzie mogą
— Największe zagrożenie: obcy mikroorganizm, który przypadkowo znajduje Ziemię idealnym środowiskiem
— Paradoks: najbardziej niebezpieczny może być najbardziej podobny do nas
Scenariusz „mutualizmu” też możliwy:
— Może obce życie wzbogaci naszą biosferę
— Nowe symbiozy, nowe szlaki metaboliczne
— Renesans biologiczny zamiast zagłady
9. OSTATECZNA ODPOWIEDŹ
Czy obce życie może zniszczyć życie na Ziemi?
TAK, ale prawdopodobieństwo zależy krytycznie od biochemicznego podobieństwa:
— Jeśli identyczne biochemicznie: wysokie ryzyko (40—60%) — konkurencja, choroby, drapieżnictwo
— Jeśli lustrzane/biochemicznie podobne: umiarkowane ryzyko (10—30%) — konkurencja o zasoby, ale nie bezpośrednia infekcja
— Jeśli zupełnie inna biochemia: niskie ryzyko (1—5%) — głównie przez zmiany środowiskowe
— Jeśli ekstremalnie różne warunki: praktycznie zerowe ryzyko — nie przeżyją na Ziemi
Najbardziej realistycznie: Przy losowym spotkaniu z obcym życiem, prawdopodobieństwo katastrofy wynosi 15—25%, ale prawdopodobieństwo całkowitego wymarcia życia na Ziemi jest niższe (5—10%) — życie jest zadziwiająco odporne, przetrwało wiele kataklizmów.
Największa lekcja:
Najniebezpieczniejsze jest to, co najbardziej podobne, ale nie identyczne — wystarczająco podobne, by wykorzystać nasze zasoby, ale wystarczająco różne, by nasze mechanizmy obronne były nieskuteczne. To właśnie dlatego programy kosmiczne mają tak ścisłe protokoły planetaryjnej ochrony — ryzyko, choć niewielkie, ma potencjalnie nieograniczone konsekwencje.
Jaki gatunek po zagładzie?
Załóżmy, że pewnego dnia znikają bezpowrotnie z Ziemi wszystkie ssaki. Pytanie: który gatunek miałby największe szanse zająć powstałą lukę biologiczną, i jakie gatunki miałyby największe szanse nająć najwyższe pozycje w szeregu ewolucyjnym (na czele którego stoi dziś człowiek)?
1. BEZPOŚREDNIE KONSEKWENCJE DLA EKOSYSTEMÓW
Ssaki zajmują kluczowe nisze:
— Megafauna roślinożerna: słonie, nosorożce, żyrafy, bizony
— Duże drapieżniki: lwy, tygrysy, wilki, niedźwiedzie
— Nocni łowcy: nietoperze (owadożerne, owocożerne, krwiopijne)
— Gryzonie: kluczowe dla rozsiewania nasion, gleby
— Naczelne: znikają wszystkie małpy, lemury
Natychmiastowe efekty:
— Eksplozja populacji owadów, gadów, ptaków (brak drapieżników)
— Zachwianie łańcuchów pokarmowych na wszystkich poziomach
— Zmiany w roślinności (brak wypasu, rozsiewania)
— Uwolnienie ogromnych zasobów (biomasa, przestrzeń)
2. KANDYDACI DO NOWEJ DOMINACJI
A. PTAKI — NAJBARDZIEJ BEZPOŚREDNI NASTĘPCY
Dlaczego ptaki?
— Najbliżsi krewni ssaków wśród kręgowców (obie grupy to amnioty)
— Już zajmują wiele nisz ssaczych (sowy = nocne drapieżniki, sępy = padlinożercy, papugi = zręczni „naczelni”)
— Wysoka inteligencja (krukowate, papugowe porównywalne z małpami)
— Zdolności manipulacyjne (dziób + łapy u niektórych)
— Komunikacja złożona (śpiew, naśladowanie)
Gatunki z największym potencjałem:
— Krukowate (Corvidae):
— Mózg względny: największy wśród ptaków (podobny do naczelnych)
— Narzędzia: wrony nowokaledońskie używają narzędzi
— Inteligencja społeczna: pamięć, planowanie, przekazywanie wiedzy
— Papugowe (Psittaciformes):
— Manipulacja: zręczne łapy + dziób
— Komunikacja: możliwość naśladowania mowy (baza dla języka?)
— Długowieczność: niektóre żyją 50—80 lat
— Sowy (Strigiformes):
— Nocne nisze po nietoperzach
— Skuteczne drapieżniki
— Ptaki bezlotne (strusie, emu, nandu):
— Mogą zająć nisze kopytnych (roślinożercy stepowi)
Potencjalna „ptasia technologia”:
— Gniazda jako „architektura”
— Narzędzia z gałęzi, kolców
— Możliwość użycia ognia? (brak rąk utrudnia)
B. GADY — POWRÓT DO ERAZ MEZOZOICZNEJ
Krokodyle i warany:
— Obecnie: ograniczone klimatem (zimnokrwistość)
— Po ociepleniu (które nastąpi bez ssaków?) — ekspansja
— Inteligencja: krokodyle mają zaskakująco rozwinięte mózgi
— Waran z Komodo: duży, społeczny, oportunistyczny
Węże i pytony:
— Zajmą nisze średnich drapieżników
— Ograniczenie: zmiennocieplność
Jaszczurki:
— Mogą wyewoluować w kierunku naczelnych jak dawniej (przodek ssaków to gady)
Potencjał ewolucyjny gadów:
— W przeszłości gady osiągały wysoką inteligencję (dinozaury ptasiomiedniczne)
— Ciepłokrwistość może się ponownie wyewoluować (tak jak u dinozaurów)
— Mózgowie gadzie ma potencjał do rozwoju
C. GŁOWONOGI — CIEMNY KONIK
Ośmiornice i kałamarnice:
— Inteligencja: już porównywalna z psami/kotami
— Manipulacja: 8 ramion z przyssawkami
— Problem: środowisko wodne ogranicza rozwój technologii
— Możliwość: kolonizacja lądu? (niektóre ośmiornice wychodzą na brzeg)
Bariery:
— Krótkie życie (1—5 lat)
— Brak społeczności wielopokoleniowej
— Rozród semelpariczny (raz w życiu)
D. OWADY — MASOWOŚĆ I SPECJALIZACJA
Termity/mrówki:
— Superorganizmy już teraz dominują wiele ekosystemów
— Rolnictwo: mrówki hodują grzyby
— Hodowla: mszyce jak bydło
— Architektura: kopce termitów
— Ograniczenie: jednostka nie jest inteligentna, tylko kolonia
Kolejne kandydaty:
— Modliszki: drapieżniki strategiczne
— Chrząszcze: różnorodność, zajmą wiele nisz
— Pszczoły: zapylacze kluczowi
E. PAJĘCZAKI I SKORUPIAKI
Pająki:
— Skuteczni drapieżnicy
— Sieci jako „technologia łowiecka”
Skorupiaki lądowe (stonogi, równonogi):
— Detrytusożercy, ważni dla gleby
3. SCENARIUSZE EWOLUCYJNE NA DŁUŻSZĄ METĘ
Skala czasowa:
10—100 lat po zniknięciu ssaków:
— Eksplozja ptaków drapieżnych i padlinożerców
— Rozrost populacji gadów w tropikach
— Owady zajmują nisze małych roślinożerców
1000—10 000 lat:
— Ptaki ewoluują w kierunku większych rozmiarów
— Nielotne ptaki zajmują nisze kopytnych
— Gady w ciepłych regionach dywersyfikują się
100 000—1 milion lat:
— Ewolucja inteligencji u krukowatych/papug
— Możliwe: narzędzia, prymitywne rolnictwo u ptaków
— Gady mogą wyewoluować ciepłokrwistość
10+ milionów lat:
— Całkiem nowe formy w zajętych niszach
— Możliwe powstanie nowej „cywilizacji”
4. KTO MA NAJWIĘKSZE SZANSE NA „INTELIGENTNĄ” DOMINACJĘ?
Ranking potencjału:
— Krukowate (45% szans)
— Już teraz: narzędzia, planowanie, kultura
— Bariera: brak rąk → ewolucja manipulacji dziobem+łapami
— Papugowe (30% szans)
— Manipulacja + komunikacja głosowa
— Długie życie, społeczność
— Gady (zwłaszcza warany/krokodyle) (15% szans)
— Jeśli wyewoluują ciepłokrwistość i większe mózgi
— Powrót do dominacji mezozoicznej
— Głowonogi (8% szans)
— Jeśli skolonizują ląd (mało prawdopodobne)
— W oceanie: rozwój podwodnych „kultur”
— Owady społeczne (2% szans)
— Inteligencja kolektywna
— Technologia oparta na feromonach/biochemii
5. NOWE „LUDZIE” — PTASI SCENARIUSZ
Potencjalna „ptasia cywilizacja”:
— Język: śpiew + gesty skrzydeł/łap
— Narzędzia: kamienie, patyki, kolce
— Rolnictwo: może uprawiać rośliny (jak niektóre mrówki)
— Ogień: trudny bez rąk, ale możliwe używanie naturalnych pożarów
— Pismo: rycie w ziemi/na korze dziobami
— Architektura: zaawansowane gniazda
Ewolucja fizyczna:
— Większe mózgi → większe głowy
— Łapy chwytne ewoluujące z obecnych
— Utrata zdolności lotu dla cięższych ciał
— Komunikacja: rozbudowany sygnał głosowy
6. NAJWIĘKSZE WYGRANE PO ZNIKNIĘCIU SSAKÓW
Natychmiastowi beneficjenci:
— Sępy i kondory — padlinożercy, tony martwych ssaków
— Duże węże (pytony, anakondy) — brak drapieżników ssaczych
— Duże ptaki drapieżne (orły, sowy) — przejmują nisze wilków, kotów
— Krokodyle/alligatory — brak konkurencji niedźwiedzi, dużych kotów
— Termity i mrówki — rozkład martwej materii, brak konkurencji gryzoni
Długoterminowi zwycięzcy:
— Ptaki z rączkami (Hoatzin ma pazurki u młodych — ewolucyjny prekursor?)
— Inteligentne ptaki (krukowate, papugi)
— Duże gady w tropikach
— Ssakopodobne gady (ewolucja w kierunku co przypominałoby ssaki)
7. CZY KTOŚ ZASTĄPI CZŁOWIEKA W PEŁNI?
Człowiek jest wyjątkowy przez:
— Ręce z przeciwstawnym kciukiem
— Mowę dzięki specyficznej budowy krtani
— Kontrolę ognia
— Kulturę kumulatywną
Żaden gatunek nie ma tego wszystkiego, ale:
Najbliżej są:
— Szympansy — ale zniknęły (są ssakami)
— Krukowate — mają kulturę, narzędzia, brakuje rąk i ognia
— Delfiny — inteligencja, ale środowisko wodne (i też ssaki)
Wniosek: Nowy dominator nie będzie kopią człowieka, ale rozwinie swoją własną ścieżkę inteligencji — może opartą na wizji, słuchu, kolektywnej inteligencji.
OSTATECZNA ODPOWIEDŹ
Największe szanse na zajęcie najwyższych pozycji ewolucyjnych:
Krótkoterminowo (100—1000 lat):
— Duże ptaki drapieżne (orły, sowy) — nisze dużych drapieżników
— Krukowate — inteligentni oportunistyczni wszystkożercy
— Duże gady w tropikach
Długoterminowo (1—10 milionów lat):
— Ptaki z rozwiniętą inteligencją (ewoluujące z krukowatych/papug) — 60% szans na nową dominującą inteligencję
— Ciepłokrwiste gady (nowe „dinozaury”) — 30% szans
— Kolektywna inteligencja owadów społecznych — 10% szans
Najbardziej prawdopodobny następca człowieka: Ptaki z rodziny krukowatych, które w ciągu 5—10 milionów lat mogą wyewoluować:
— Większe mózgi
— Lepsze manipulacje (ewolucja łap)
— Kompleksową komunikację głosową
— Prymitywną technologię (narzędzia, może kontrola ognia)
Ziemia bez ssaków stałaby się planetą ptaków z wyspami gadziej dominacji w tropikach, a nowa „cywilizacja” byłaby ptasią, nie ssaczą.
Implant w mózgu?
Gdyby jakiejś niezbyt odległej przyszłość wszyscy żyjący ludzie zostali „zaczipowani” (wszczepiono by im chipy elektroniczne do głów czy też do innych organów,) jak zmieniłoby się życie?
Era Neuroimplantów: Kiedy Człowiek Staje Się Cyborgiem
WPROWADZENIE: PRZEŁOMOWY MOMENT W EWOLUCJI GATUNKU
Wizja powszechnych implantów neuronalnych przestaje być domeną science fiction, stając się technologiczną nieuniknionością. Gdyby w niezbyt odległej przyszłości — powiedzmy 2045—2070 roku — każdy człowiek otrzymał neuroimplant, byłaby to największa transformacja w historii gatunku ludzkiego, porównywalna jedynie z odkryciem ognia, wynalezieniem pisma czy rewolucją cyfrową. Nie chodzi tu o proste identyfikatory czy płatnicze chipy podskórne, lecz o zaawansowane interfejsy mózg-komputer (BCI), które bezpośrednio łączą biologiczną świadomość ze światem cyfrowym. Taki scenariusz rodzi fundamentalne pytania: czy wciąż bylibyśmy ludźmi w tradycyjnym sensie? Jak zmieniłyby się relacje społeczne, gospodarka, polityka, religia? Czy powstałaby nowa forma świadomości — hybryda biologiczno-cyfrowa? Niniejsza analiza ma na celu szczegółowe zbadanie konsekwencji powszechnej „chipizacji” ludzkości, rozważając zarówno utopijne wizje wzmocnienia człowieka, jak i dystopijne scenariusze totalitarnej kontroli.
ROZDZIAŁ 1: TECHNOLOGICZNE PODSTAWY — JAK DZIAŁAŁBY UNIWERSALNY NEUROIMPLANT?
1.1. Architektura systemu
Uniwersalny neuroimplant przyszłości prawdopodobnie składałby się z:
1.1.1. Komponent fizyczny:
— Neuronalna siatka (neural lace) — elastyczna, biokompatybilna sieć mikroelektrod pokrywająca korę mózgową
— Procesor neurosynaptyczny — specjalizowany chip przetwarzający sygnały neuronalne
— Moduł komunikacyjny — 5G/6G/7G, Li-Fi, czy komunikacja kwantowa
— Zasilanie — harvesting energii z glukozy we krwi, ruchu ciała, różnic temperatur
— Pamięć nieulotna — nanowłókna DNA lub pamięć atomowa o pojemności petabajtów
1.1.2. Warstwa oprogramowania:
— System operacyjny świadomości (Consciousness OS) — zarządzanie zasobami neuronalnymi
— API neuronowe — standardy komunikacji między mózgiem a urządzeniami zewnętrznymi
— Wirtualne asystenty — AI zintegrowane z procesami myślowymi
— Zabezpieczenia — neurokryptografia, firewalle mentalne
1.1.3. Infrastruktura zewnętrzna:
— Neurochmura (NeuroCloud) — rozproszona sieć serwerów przetwarzających dane neuronalne
— Stacje bazowe — zapewniające zasięg neurokomunikacji
— Centra diagnostyczne — monitoring zdrowia neuronalnego
— Archiva pamięci — zdalne kopie zapasowe świadomości
1.2. Podstawowe funkcjonalności
1.2.1. Komunikacja bezsłowna:
Kup książkę, aby przeczytać do końca.